גידי נאמן החוג לביולוגיה וסביבה, אוניברסיטת חיפה-אורנים gneeman@gmail.com
עדו יצחקי החוג לביולוגיה אבולוציונית וסביבתית, אוניברסיטת חיפה izhaki@research.haifa.ac.il
תקציר: מחקר זה מתאר את הסוקצסיה בחלקות שונות של כרם אחד גדול, שננטשו במועדים שונים. התוצאות מתארות את שלבי ההתנחלות של צמחי-בר רב-שנתיים בחלקות הנטושות. המחקר מנתח את הגורמים ובעיקר את תכונות הצמחים המשפיעים על הרכב המינים המתנחלים ומשווה את התוצאות לתאוריה הקלאסית של הסוקצסיה בשדות נטושים וסוקצסיה בכרמים נטושים במערב הים-התיכון. מאמר זה מתבסס על המאמר: Ne’eman, G. and Izhaki, I. 1996. Colonization in an abandoned East Mediterranean vineyard. Journal of Vegetation Science 7: 465-472.
————————-
תקציר: השתקמות הטבעית של צמחי בר לאחר הפרעה נקראת סוקצסיה. על פי התאוריה הקלאסית הסוקצסיה מתחילה במיני חלוץ, מצורות חיים נמוכות (עשבוניים חד-שנתיים, עשבוניים רב-שנתיים ובני-שיח), אשר בנוכחותם מטייבים את תנאי הגידול בשדה, בעיקר את הקרקע, ומאפשרים בכך את התבססות של מינים אחרים מצורות חיים גבוהות יותר (שיחים ועצים) אשר לאחר התבססותם דוחקים את מיני החלוץ ושולטים בשטח עד להפרעה הבאה (כגון כריתה, רעיה, שריפה), או שהם מוחלפים על-ידי מיני-קיימא שהם בדרך כלל עצים השולטים בשטח במצב של העדר הפרעה ומרכיבים את צומח השיא (קליימקס) באזור. צומח השיא מושפע בעיקר מתנאי האקלים והמסלע. מחקרים קודמים שעסקו בדינמיקה של התנחלות צמחי בר בשדות וכרמים נטושים נערכו במערב אגן הים התיכון, אבל בישראל כמעט ולא נאספו נתוני שדה בנושא הסוקצסיה. כאן אנו מציגים את הסוקצסיה של צמחים רב-שנתיים בחלקות שונות של כרם אחיד שננטשו 5, 8, 15, ו- 35 שנים לפני תחילת המחקר שנערך בחורף 1987-8, ומאז לא סבלו מהפרעה ניכרת. בניגוד לתאוריה, מיני הצמחים הראשונים שהתנחלו בכרם לא היו של צורות חיים נמוכות, אלא מינים רב-שנתיים מעוצים שכולם בעלי פירות בשרניים שזרעיהם הופצו לכרם כנראה על-ידי ציפורים מהחורש הטבעי הסמוך. הגפנים הנטושים ועמודי ההדליה היוו מקומות עמידה ותזונה לציפורים ובכך זרזו את ההגעה של מיני צמחים שזרעיהם מופצים על-ידי צפרים, זאת ללא תלות בצורת החיים שלהם. בתנאים אלה דרך הפצת הזרעים ולא צורת החיים הייתה התכונה העיקרית שהשפיעה על קצב התנחלות. מכיוון שבין צמחי החלוץ היו גם עצים ניתן להסיק שתנאי בית הגידול בכרם הנטוש היו טובים ולא נדרשו שלבים מקדימים לשיפור תנאי בית הגידול ולא נצפתה החלפה של מינים במשך השנים, כפי שמקובל בתאוריה הקלאסית. קצב הסוקצסיה היה מהיר ונדרשו רק כ 8 -15 שנים להתייצבות הרכב המינים, לצמיחה לגובה הרגיל של העצים ולהשגת אחוז כיסוי גבוה של המינים המתנחלים.
=====================
מבוא
סוקצסיה היא התהליך בו משתקמת צמחייה טבעית לאחר הפרעה. סוקצסיה ראשונית מתרחשת כאשר השטח נקי מכל צמחים כמו באי חדש העולה מן האוקיינוס, או שטח שכוסה לבה בזמן התפרצות געשית. אולם במקרים בהם ההפרעה אינה מחלטת ובשטח נשארו צמחים חיים או זרעים שלהם בקרקע, מכונה התהליך סוקצסיה משנית. מחקרים רבים נערכו על הסוקצסיה המשנית לאחר ששדות רבים ננטשו בצפון מזרח ארה"ב בעקבות הבהלה לזהב, וצמחי בר התנחלו בהם (Billings, 1938; Oosting, 1942; Buel et al., 1971; Pickett, 1982; Gill & Marks, 1991).
התאוריות הראשונות של הסוקצסיה ואלו שבאו בעקבותיהן (Clements, 1904, 1916; Odum, 1969) היו הוליסטיות והניחו שהשינוי בהרכב הצמחייה הוא דטרמיניסטי ונקבע על פי מצב השטח לאחר ההפרעה וצומח השיא (קליימקס) במקום נקבע בעיקר על פי תנאי האקלים והמסלע. על פי תאוריות אלה, הכוחות המניעים הם תכונה של המערכת האקולוגית והשינויים בהרכב חברות הצמחים הם תוצאה של יחסים בין המינים בחברת הצמחים בכל זמן נתון. תאוריות מאוחרות יותר הציעו שהשינוי בחברה הוא תוצאה של יחסים בין אוכלוסיות בתנאי סביבה משתנים. התאוריות המאוחרות מציגות הסברים מכניסטיים לסוקצסיה בתנאי סביבה משתנים והעדר שווי משקל יציב בין הצמחים לסביבה (Glenn-Lewin et al., 1992).
המנגנונים הקובעים את השנויים בהרכב המינים במשך הסוקצסיה נידונו בהרחבה בספרות המדעית (למשל: Drury & Nisbet, 1973; Connell & Slatyer, 1977; Horn 1981; Glenn-Lewin & van der Maarel, 1992; van Andel et al., 1993), ועיקרם הם:
- הפצת זרעים וטריפתם, נביטה וטריפת נבטים, תחרות בין נבטים וסיכויי ההתבססות שלהם (Gill & Marks, 1991).
- הרכב המינים ההתחלתי ('Initial floristic composition') והיתרון היחסי של מיני החלוץ המתנחלים תחילה ('pre-emption') לפי (Eglerת 1954).
- התכונות המובנות של צורות החיים (עשבוני, בן שיח, שיח ועץ) (Pickett, 1976; Drury & Nisbet, 1973) והתכונות האקופיסיולוגיות שלהם (Bazzaz, 1979).
- יחסי הדדיות בין המינים כולל: (1) תחרות על משאבי הסביבה המשתנים במשך התהליך (Tilman, 1987, 1988). (2) תגובה (reaction – Clements, 1916) או עידוד (facilitation- Connell & Slatyer, 1977), כאשר צמחי חלוץ משנים את תנאי הסביבה ובכך מאפשרים התנחלות של מינים נוספים. (3) עיכוב, כאשר מינים קיימים מונעים נביטה צמיחה או התבססות של מינים אחרים (Connell & Slatyer, 1977). (4) סובלנות, כאשר אין עיכוב או עידוד (Connell & Slatyer, 1977).
ניסויים ומחקרים מאוחרים יותר הראו שרק פעולה משולבת של מספר מנגנונים יכול להסביר את הסוקצסיה במערכות אקולוגיות (Gill & Marks, 1991; Chapin et al., 1994). במחקרים שנערכו במערכות אקולוגיות שונות במקומות שונים בעולם, נמצאו הבדלים בולטים במנגנונים המניעים את הסוקצסיה. על מנת לקדם את הפיתוח של תאוריה כוללנית המסבירה את הגורמים והמנגנונים השונים לסוקצסיה יש חשיבות רבה לאסוף כל מידע אפשרי על סוקצסיה במערכות אקולוגיות שונות (van Andel et al., 1993; McCook, 1994).
במקרים רבים, חקר הסוקצסיה עוקב אחרי התפתחות הצומח והצמחייה בחלקות קבועות, מחקר ארוך טווח שנמשך שנים רבות. שיטה מחקר נוספת מתבססת על סדרת זמן (chronosequence) = (Glenn-Lewin & van der Maarel, 1992), או כפי שהיא מכונה 'התמרה של מקום בזמן' (space for time substitution) (Pickett, 1989). בשיטה זו מאתרים חלקות שהיו דומות מאוד לפני ההפרעה אך הן הנבדלות בפרקי הזמן שעברו מאז ההפרעה האחרונה. הביקורת העיקרית לגבי שיטה זו היא שחלקות שונות אינן חזרות אמתיות שכן הן עשויות להיות שונות מעט אלה מאלה בהרכב הקרקע, מיקרו אקלים, הפרעות קלות אחרות וזמינות של זרעים של מינים מתנחלים (Pickett, 1989), או הבדלים אחרים שהחוקר אינו מודע להם.
מחקרים כמותיים נערכו על סוקצסיה במטעים נטושים בוצעו בספרד ודרום צרפת (,Houssard et al., 1980; Debussche et al., 1982; Debussche et al., 1985; Escarre et al. 1983; Debussche & Lepart, 1992; Debussche & Isenmann, 1994; Tatoni & Roche, 1994). למיטב ידיעתנו לא בוצעו מחקרים דומים בישראל.
המחקר הנוכחי נערך בשיטת סדרת הזמן בחלקות דומות של כרם גדול שננטשו בזמנים שונים. מטרות המחקר היו: (1) לנטר את הרכב המינים הרב-שנתיים. (2) לנתח את הרכב המינים על פי צורות החיים ודרכי הפצת הזרעים שלהם. (3) להעריך את משך הזמן הדרוש לצמיחה ולכיסוי מלא על-ידי צמחים רב-שנתיים. (4) להשוות את קצב הסוקצסיה בין צמחי הגפן בתוך השורות לזו המתרחשת בין השורות של הגפנים הנטושות. ו (5) להשוות את הסוקצסיה בכרם נטוש בישראל לזו המתרחשת בחלקו המערבי של אגן הים התיכון.
שיטות
שטח המחקר
המחקר נערך בכרם בשטח של כעשרה דונמים שניטע ב 1958 במושב אמירים (35° 56' E, 32° 56' N) הנמצא בגבול בין הגליל התחתון לגליל העליון. הצמחים ניטעו במרחקים של שני מטרים זה מזה בתוך השורה ורווח של שני מטרים בין השורות. לאחר הנטיעה, חולק הכרם לחלקות של 400-300 מ"ר שעובדו על-ידי משפחות שונות. הדליית הגפנים הייתה בעמודי ברזל בגובה של מטר וחצי, הכרם הושקה בקיץ, קולטר כפעמיים בשנה בין השורות ולא רוסס נגד עשבייה. בשל סיבות סוציואקונומיות ננטשו חלקות הכרם בהן נערך המחקר חמש, שמונה, 15 ו- 35 שנים לפני תחילתו (המידע מבעלי הכרמים). מאז נטישת הכרם ועד עריכת המחקר באביב 1993, לא הייתה במקום הפרעה משמעותית כגון שריפה או רעייה. עם נטישת הכרם הופסקו ההשקיה והקילטור, אך הגפנים ועמודי ההדליה לא נעקרו.


מימין – מיקום חלקת המחקר בכרם אמירים, כיום – שטח מרעה; משמאל – כרם נטוש (אינו הכרם שבו נערך המחקר)
לצפייה מיטבית – לחצו על התמונות


בכל חלקה היו 10 שורות, על מנת להמנע מאפקט שוליים, נדגמו השורות 6,4,2, ו- 8. בתוך כל שורה נדגמו חמישה ריבועים כל אחד של מ"ר אחד, כשבמרכז הריבוע הייתה גפן. נדגמו גם חמישה ריבועים כל אחד של מ"ר אחד ברווח שבין השורות, במרחק המירבי האפשרי משתילי הגפנים. אורך השורות היה כ 50 מ' והמרחק בין הריבועים היה כ 10 מטרים. בסך הכול נדגמו בכל חלקה 20 ריבועים בתוך השורות ו 20 בין השורות.
הכרם נמצא במפנה דרומי מתון מאוד ברום של 180 – 260 מטרים מעל לפני הים. האקלים ים-תיכוני טיפוסי עם 654 מ"מ גשם ממוצע בשנה וטמפרטורות ממוצעות של 9.2 ו 26.1 מעלות צלזיוס בינואר ואוגוסט, בהתאמה. הקרקע היא טרה רוסה קלציטית (רבינוביץ-וין, 1986) רדודה (30 ס"מ) בעלת תגובה (pH) בסיסית מעט (8.0-7.9) שנמדדה בזמן הנטיעה. בשל הטופוגרפיה המתונה והקרבה הרבה של החלקות ניתן להניח שהתנאים הסביבתיים בכל החלקות דומים מאוד.
הכרם היה מוקף בחברת חרוב מצוי ואלת המסטיק (זהרי, 1959) עם אלה ארץ-ישראלית, אלון מצוי ואלון התבור כמינים מלווים. סירה קוצנית וקידה שעירה נפוצים מאוד במרווחים שבין עצי החורש. המרחק המרבי בין מרכז הכרם לשוליו היה כ 100 מ' ורק כ 40 מ' בין השוליים לכל אחת מחלקות המחקר. מכיוון שמרחק הפצת הזרעים על-ידי ציפורים באגן הים התיכון הוא כ 100 מ' (Debussche & Isenmann, 1994), ניתן להניח שלא היו הבדלים בזמינות זרעים מופצי ציפורים בין החלקות.
להשוואת הממוצעים בין השנים השתמשנו במבחן Kruskal-Wallis, להלן בהמשך (K.W).
תוצאות
אחוז כיסוי
בתוך השורות של הכרם היה אחוז הכיסוי של הצמחים המעוצים הרב-שנתיים גבוה מאוד (53%) כבר חמש שנים לאחר הנטישה, בעוד שבין השורות הוא הגיע לשיעור דומה רק 35 שנים לאחר הנטישה. אחוז הכיסוי של המעוצים בתוך השורות התייצב על 90-80% כבר שמונה שנים לאחר הנטישה (טבלה 1). אחוז הכיסוי של העשבוניים החד-שנתיים היה בעל מתאם שלילי עם אחוז הכיסוי של המעוצים. בתוך השורות אחוז הכיסוי שלהם החל לרדת באופן חד לאחר שמונה שנים עד להיעלמותם המחלטת (איור 1). בניגוד לכך בין השורות אחוז הכיסוי שלהם היה גבוה (>70%) חמש עד 15 שנים לאחר הנטישה, עם ירידה חדה לאחר מכן. אחוז הקרקע החשופה היה תמיד גבוה יותר בין בשורות מאשר בתוכן (טבלה 1).
טבלה 1. אחוז הכיסוי (ממוצע±סטיית תקן) של צמחים עשבוניים חד-שנתיים, רב-שנתיים מעוצים וקרקע חשופה בתוך השורות (n=20)ובין השורות (n=20) בחלקות של הכרם: חמש, שמונה, 15 ו-35 שנים לאחר הנטישה.
| שנים מאז הנטישה | 5 | 8 | 15 | 35 |
| בתוך השורות | ||||
| עשבוניים | 45±22 | 7±4 | 15±22 | 0 |
| מעוצים | 53±24 | 91±8 | 82±21 | 82±31 |
| קרקע חשופה | 5±3 | 4±3 | 6±2 | 25±28 |
| בין השורות | ||||
| עשבוניים | 89±12 | 72+32 | 74±34 | 18±27 |
| מעוצים | 0 | 22±33 | 21±35 | 50±35 |
| קרקע חשופה | 11±12 | 10±17 | 6±3 | 37±34 |
נמצאו הבדלים מובהקים באחוז הכיסוי של הצמחים המעוצים הרב-שנתיים (.χ 2=28.34, df=3, P<0.05, K.W), העשבוניים החד-שנתיים (.χ2= 58.12, df=3, P<0.05, K.W) והקרקע החשופה (.χ2=25.45, df=3, P<0.05, K.W) בתוך השורות בין השנים מאז הנטישה. ההבדלים באחוז הכיסוי של המעוצים הרב-שנתיים (.χ 2=33.78, df=3, P<0.05, K.W), העשבוניים החד-שנתיים (.χ 2=36.65, df=3, P<0.0005, K.W) והקרקע החשופה (.χ 2=17.91, df=3, P<0.05, K.W) בין השנים מאז הנטישה בין השורות היו מובהקים גם הם.
גובה הצמחים המעוצים הדומיננטיים
גובה הצמחים המעוצים הדומיננטיים של אלה ארץ-ישראלית, אלת המסטיק ואשחר רחב עלים שצמחו בתוך השורות בחלקות חמש שנים לאחר הנטישה, לא היה שונה מגובהם בחלקות 35 שנים לאחר מכן (Wilcoxon rank-sum two sample test) – לאלה ארץ-ישראלית (Z=-0.85, P>0.1), לאלת המסטיק (Z=-0.55, P>0.10) ולאשחר רחב עלים (Z=-0.24, P>0.10). כבר חמש שנים לאחר הנטישה הגיעו שלושת המינים לגובהם הרגיל וגבהו רק מעט במשך 30 השנים שלאחר מכן (טבלה 2).
טבלה 2. הגובה (ס"מ) הממוצע (±סטיית תקן) של הצמחים של אלה ארץ ישראלית, אלת המסטיק ואשחר רחב עלים שגדלו בתוך שורות הגפנים בחלקות חמש שמונה, 15 ו 35 שנים מאז הנטישה (n=20).
| שנים מאז הנטישה | 5 | 8 | 15 | 35 |
| אלה ארץ ישראלית | 46±165 | 55±105 | 37±227 | 66±204 |
| אשחר רחב עלים | 42±180 | 124±138 | 103±215 | |
| אלת המסטיק | 42±70 | 28±58 | 103±215 | 44±94 |
עושר המינים וקצב ההתנחלות של מינים מעוצים רב-שנתיים
השינוי העיקרי בצפיפות הצמחים הרב-שנתיים (מספר הפרטים למ"ר) חל בין חמש לשמונה שנים לאחר נטישת הכרם, כאשר הצפיפות עלתה פי ארבע בתוך השורות, ובפרק הזמן שבין 15 ל 35 שנים לאחר הנטישה (טבלה 3). צפיפות הצמחים המרבית הושגה כבר שמונה שנים לאחר נטישת הכרם וההבדלים בין שמונה ל 35 שנים מהנטישה לא היו מובהקים (Wilcoxon two sample test, Z=0.6, P>0.5). תוצאות דומות התקבלו לגבי צפיפות המינים (מספר מינים למ"ר). צפיפות המינים המרבית הושגה כבר שמונה שנים לאחר הנטישה (טבלה 3) ולא השתנתה באופן מובהק גם לאחר 35 שנים (Wilcoxon two sample test, Z =-0.78, P>0.1). צפיפות המינים בין השורות הייתה נמוכה בהרבה ורק 35 שנים לאחר נטישת הכרם השתוותה לצפיפות המינים בתוך השורות (טבלה 3).
עושר המינים (מספר המינים הרב שנתיים הכללי לחלקה) הרב-שנתיים בתוך השורות הגיע ל 15 כבר שמונה שנים לאחר נטישת הכרם ונשאר יחסית קבוע בשנים שלאחר מכן. מספר המינים החדשים שהתנחלו בתוך השורות ירד באופן חד מ 15 מינים בחמש השנים הראשונות לאחר נטישת הכרם למין אחד בלבד בין 15 ל 25 שנים לאחר הנטישה.
טבלה 3. צפיפות הצמחים (מספר פרטים למ"ר) הממוצעת (±סטיית תקן) וצפיפות המינים (מספר המינים למ"ר) הממוצעת (±סטיית תקן) של הצמחים המעוצים הרב-שנתיים בתוך השורות (n=20) וביניהן (n=20) חמש שמונה, 15 ו 35 שנים לאחר הנטישה.
| שנים מאז נטישת הכרם | 5 | 8 | 15 | 35 |
| בתוך השורות | ||||
| צפיפות פרטים | 2.7±2.0 | 9.8±4.9 | 6.7±2.2 | 8.6±3.7 |
| צפיפות מינים | 1.9±1.1 | 3.6±0.8 | 2.4±1.0 | 4.1±1.7 |
| בין השורות | ||||
| צפיפות פרטים | 0 | 1.5±1.6 | 0.9±1.4 | 5.5±5.0 |
| צפיפות מינים | 0 | 1.1±1.0 | 0.7±1.1 | 2.3±0.9 |
צורות החיים
כל מיני הצמחים הרב שנתיים שנמצאו בכל החלקות סווגו על פי צורות החיים שלהם (פיינברון ודנין 1990) (נספח 1). עצים ומטפסים היו צורות החיים הראשונות שהתנחלו בכרם הנטוש ועשו זאת תחילה בתוך שורות הגפנים (איור 1).
בתוך שורות הגפנים לא נמצאו הבדלים מובהקים בצפיפות מיני (מספר המינים הממוצע במ"ר) העצים והמטפסים בין החלקות שננטשו חמש, שמונה, 15 ו35 שנים לפני המחקר (K.W. χ 2=3.699, df=3, P>0.05,). צפיפות מיני השיחים הייתה נמוכה תחילה אבל עלתה באופן מובהק (.χ2=21.59, df=3, P<0.01, K.W). משך השנים (איור 1). צפיפות מיני בני-השיח ועשבוניים רב-שנתיים הייתה נמוכה מאוד בכל החלקות (איור 1).
בין השורות הייתה צפיפות מיני השיחים הגדול ביותר וגדל באופן מובהק עם השנים (.χ2=28.54, df=3, P<0.01, K.W). רק לאחר 35 שנים השתוותה צפיפות מיני השיחים בין השורות לזו שהייתה בתוך השורות חמש שנים לאחר נטישת הכרם (איור 1).

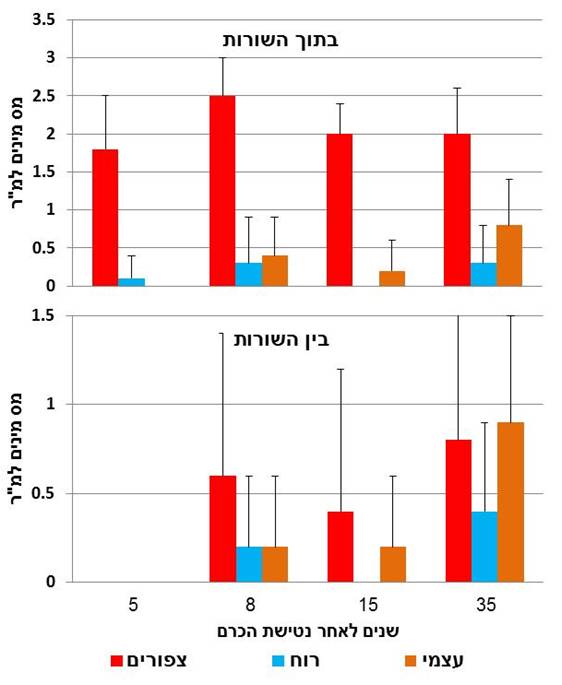
איור 1 (מימין). התפלגות מספר מיני הצמחים הרב-שנתיים (צפיפות המינים הממוצעת + סטיית תקן) על פי צורות החיים שלהם בתוך השורות (n=20) ובין השורות (n=20) של הכרם חמש, שמונה, 15 ו 35 שנים לאחר הנטישה. רשימת המינים וצורת החיים של כל אחד מהם ראו נספח 1 בסוף המאמר.
איור 2 (משמאל). התפלגות מספר מיני הצמחים הרב-שנתיים (צפיפות המינים הממוצעת + סטית תקן) על פי צורות הפצת הזרעים שלהם (יצחקי 1986) בתוך השורות (n=20) ובין השורות (n=20) של הכרם חמש, שמונה, 15 ו-35 שנים לאחר הנטישה. רשימת המינים וצורת החיים של כל אחד מהם ראו נספח 1 בסוף המאמר.
להגדלה – לחצו על התמונות
צורות הפצה
מיני הצמחים חולקו על פי צורות הפצת הזרעים שלהם: ברוח, עצמית או על-ידי צפרי שיר. אופן ההפצה וציפורי השיר הידועות כמפיצות הזרעים (יצחקי, 1986) מוצגים בנספח 1 (ראו בסוף המאמר).
בתוך שורות הגפנים הייתה הצפיפות של מיני הצמחים הרב-שנתיים (מספר מינים למ"ר) שזרעיהם מופצים על-ידי ציפורי שיר גבוהה יותר מזו של המינים שזרעיהם מופצים ברוח או עצמית בכל השנים שלאחר נטישת הכרם (איור 2) וההבדל בין השנים היה מובהק (χ2=10.96, df=3, P<0.05, K.W). מינים שזרעיהם מופצים ברוח היו נוכחים בחלקות כבר חמש שנים לאחר הנטישה בעוד שזמינים שזרעיהם מופצים עצמית התנחלו בכרם הנטוש רק בשלב מאוחר יותר (איור 2). ההבדלים בצפיפות המינים בין השנים הייתה מובהקת במינים שזרעיהם מופצים ברוח (χ2=9.98, df=3, P<0.05, K.W) ובאופן עצמי (χ2=23.57, df=3, K.W P<0.001).
בין שורות הגפנים בכרם הנטוש נמצאו צמחים רב-שנתיים רק החל משמונה שנים לאחר הנטישה (איור 2). גם כאן היו הבדלים מובהקים בין השנים בצפיפות המינים שזרעיהם מופצים על-ידי ציפורים (χ2=16.54, df=3, P<0.01, K.W) רוח (χ2=22.91, df=3, P<0.01, K.W) ועצמית (χ2=35.56, df=3, P<0.01, K.W), אבל לא הייתה מגמה שינוי ברורה עם שנים (איור 2).
דיון
החלפת מינים
התאוריה הקלאסית של הסוקצסיה מדגישה את תהליך החלפת המינים עם חלוף הזמן מאז ההפרעה (Clements, 1916) וחלוקה של המינים למיני חלוץ ומיני קיימא המחליפים אותם (Bazzaz, 1979). באתר המחקר שלנו צמחים חד-שנתיים יכולים להיחשב כמיני חלוץ, כפי שנמצא גם בכרמים נטושים במערב אגן הים-התיכון (,Houssard et al., 1980; Escarre et al. 1983). בשלבים הראשונים של הסוקצסיה צמחים חד-שנתיים היו נפוצים בין שורות הגפנים. הירידה באחוז הכיסוי שלהם והיעלמותם מתוך השורות לאחר 35 שנות נטישה של הכרם (טבלה 1) מתאימים לתאוריית ההחלפה של מיני החלוץ. אולם קרוב לוודאי שמינים חד-שנתיים אלו היו בכרם כעשבים רעים עוד טרם הנטישה וזרעיהם היו בבנק הזרעים בקרקע.
אלת המסטיק ומיני הצמחים המטפסים סווגו כצמחי קיימא (קליימקס) של החורש הים-תיכוני (זהרי, 1959), אבל הם כבר נמצאו בחלקות הכרם כבר חמש שנים לאחר הנטישה. יתרה מזאת מיני עצים כמו אלה ארץ–ישראלית ואשחר רחב-עלים השיגו את גובהם היציב כבר 8 -15 שנים לאחר נטישת הכרם (טבלה 2) בדומה לגובהם בשטחי חורש סמוכים ולכן גם לא נראה סביר שיוחלפו בעתיד הנראה לעין על-ידי מינים אחרים. בשל העדר כמעט מוחלט של החלפת המינים בחלקות הכרם הנטוש, עושר המינים הרב-שנתיים וצפיפותם התייצבו כבר שמונה שנים לאחר הנטישה (טבלה 3). מצב זה בו מיני קיימא מתבססים בשטח זמן קצר לאחר הנטישה, שונה מהדינמיקה בכרמים נטושים במערב אגן הים-התיכון שם תוארה סדרה קלאסית של החלפת מינים: חד-שנתיים הוחלפו על-ידי מינים ממשפחת השפתניים ואחריהם דגניים רב-שנתיים שהוחלפו על-ידי מיני שיחים שזרעיהם מופצים על-ידי ציפורים ומינים של אלונים (Houssard et al., 1980; Escarre et al., 1983).
בכרם הנטוש באמירים לא ניתן לחלק את המינים הרב-שנתיים למיני חלוץ ומיני קיימא. התנאים הסביבתיים ובמיוחד תכונות הקרקע בכרם היו טובים דיים להתבססות מיני הקיימא ולא היה צורך בשיפור התנאים על-ידי מיני חלוץ כפי שמקובל בתאוריה הקלאסית (Connell & Slatyer, 1977). מכיוון שכמעט כל המינים שהתנחלו בכרם נשארו שם לפחות למשך 35 שנים ניתן להניח שגם לא הייתה דחיקה תחרותית של מינים כפי שמתואר בתאוריה הקלאסית (Tilman, 1987).
צורת החיים ודרך הפצת הזרעים
התאוריה המקובלת של הסוקצסיה של צמחים בשדות נטושים באזור הים-תיכוני מבוססת על החלפה מיני הצמחים על פי צורת החיים שלהם. מיני החלוץ הם מינים חד-שנתיים המוחלפים על-ידי בני-שיח ליצירת בתה, לאחר מכן מחליפים שיחים את בני-השיח ליצירת גריגה ורק אז מתנחלים מיני עצים שונים ליצירת החורש הים-תיכוני המייצג את צומח השיא בהעדר הפרעה (זהרי, 1959). דגם דומה של סוקצסיה תואר גם בכרמים נטושים בדרום צרפת (Houssard et al., 1980; Escarre et al., 1983). כאמור בניגוד לשם, בכרם הנטוש באמירים, מרבית המינים הראשונים שהתנחלו בתוך שורות הגפנים היו מיני עצים ומטפסים (איור 1) ולא בני-שיח ועשבוניים רב-שנתיים, בדומה לתיאור הסוקצסיה אצל חריף (1974) ורבינוביץ-וין (1986). אולם יש לזכור שתוצאות מחקר זה משקפות את המצב בכרם נטוש אחד ולכן עדיין לא ניתן להכליל אותם לכרמים ושדות נטושים אחרים.
ההתנחלות המהירה יותר של עצים ומטפסים בתוך שורות הגפנים בכרם הנטוש באמירים מאשר ביניהם, לא יכולה להיות מוסברת על-ידי צורות החיים שלהם (איור 1), אבל ניתן להסבירה על-פי דרך הפצת הזרעים שלהם שהיא באמצעות ציפורים האוכלות את הפירות ומפרישות את הזרעים (אנדוזואוכוריה). דרך הפצת הזרעים של מיני צמחים תוארה כתכונת מפתח חשובה ביותר במהלך הסוקצסיה (Noble & Slatyer, 1980). גם בכרם הנטוש באמירים היה אופן הפצת הזרעים המכניזם המסביר באופן הטוב ביותר את סדר ההתנחלות של המינים. מספר המינים למ"ר של מיני צמחים שזרעיהם מופצים על-ידי ציפורים היה הגבוה ביותר בעיקר בתוך שורות הגפנים (איור 2) ללא תלות בצורת החיים שלהם (איור 1). אחוז המינים שזרעיהם מפוצים על-ידי ציפורים בחורש הים תיכוני בישראל הוא 24% (יצחקי 1986) בעוד שבכרם הנטוש באמירים שיעורם היה 58% (נספח 1). ייצוג יתר זה מלמד שהגעת המינים לכרם הנטוש לא הייתה אקראית ושלציפורים יש תפקיד חשוב בתהליך זה. יש להניח שעוד לפני נטישת הכרם נמשכו ציפורים אוכלות פירות לענבים הבשלים בכרם וגם לאחר הנטישה היו צמחי הגפן שלא נעקרו ועמודי ההדליה מקומות ישיבה מועדפים לציפורים שניזונו בפירות צמחי החורש והפרישו חלק מהם בתוך שורות הגפנים בכרם אחרי הנטישה וכנראה גם לפני כן. מטפסים ושיחים שזרעיהם נפוצים על-ידי ציפורים גדלים באופן קבוע סמוך לגזעי עצים במטעים שונים בגליל (מידע אישי).
לצמחים בולטים בשטח ולמקומות עמידה מלאכותיים נמצאה השפעה חיובית על קצב הסוקצסיה והיא תועדה רבות בשדות נטושים בצפון מזרח ארה"ב (McDonnell & Stile,s 1983; Holthuijzen & Sharik, 1985; McClanahan & Wolfe, 1993), בשדות מרעה במקסיקו (Guevara et al. 1986; 1992) ובמטעים נטושים בדרום אירופה (Houssard et al. 1980; Debussche et al., 1982; Debussche & Lepart, 1992; Escarre et al., 1983; Debussche & Isenmann, 1994). לציפורים אוכלות פירות ומפיצות זרעים עשויה להיות השפעה על עיצוב הצמחייה, אבל לצמחייה הקיימת השפעה על גיוס והתנחלות של צמחים חדשים באמצעות השפעתם על דגם התנועה של ציפורים ועל הזרעים המופצים על ידם (McDonnell & Stiles, 1983; McClanahan & Wolfe, 1993). הצבת מקומות עמידה לציפורים מפיצות זרעים נהוגה היום כשיטה מקובלת לשיקום נופי צומח פגועים באגן הים התיכון (,Pausas et al. 2004) ובמקומות שונים בעולם (למשל: Holl, 1998; Holl et al., 2000).
בהשוואת תהליך הסוקצסיה בין ישראל לצרפת עולה כי הצמחים שזרעיהם מופצים על-ידי ציפורים היו הראשונות להתנחל בישראל ורק במקום הרביעי בצרפת, כמו כן בישראל אחרי 15 שנים הייתה התייצבות בהרכב המינים ובגובה של העצים בעוד שבצרפת תהליך זה נמשך כ-60 שנים (Escarre et al., 1983). ההבדל בקצב הסוקצסיה יכול להיות מוסבר בכך שהכרמים בצרפת היו מרוחקים משטחי חורש טבעי ועם הנטישה הם נעקרו (Escarre et al,. 1983). לעומת זאת הכרם באמירים שהיה מוקף בחורש לא נעקר בעת הנטישה. בנוסף לכך בצרפת גידול הכרמים נמשך שנים רבות באותם השטחים ובאמירים הנטיעה הייתה בשטח חדש שהוכשר וידוע שקצב הסוקצסיה נמצא ביחס הפוך למשך העיבוד החקלאי במקום (Myster & Pickett 1994). אין לנו הסבר טוב לקצב המהיר יותר של הסוקצסיה בישראל מאשר בצרפת, במיוחד לאור העובדה שכמות המשקעים בצרפת (1200 מ"מ בשנה) הייתה כמעט כפולה מזו שבישראל (כ-650 מ"מ בשנה). אולם יתכן שמחסור בזרעים של צמחי קיימא עיכב את התהליך בצרפת יחסית לכרם אמירים שהיה סמוך מאוד למקור הזרעים בחורש. לסיכום, הרכב המינים של הציפורים אוכלות הפירות ומפיצות הזרעים דומה בישראל ובצרפת ובשני המקרים נמצא דמיון בהשפעה של נוכחות מיני הציפורים, מיני צמחים שזרעיהם מופצים על-ידי ציפורים על הדינמיקה של הסוקצסיה המשנית בכרמים נטושים.
ההשפעה המכרעת של הפצת זרעים על-ידי ציפורים בכרם הנטוש מודגשת בכך ששני מינים הנפוצים מאוד בחורש שסבב את הכרם (חרוב מצוי ואלון מצוי) אבל זרעיהם אינם מופצים על-ידי ציפורים אוכלי פירות לא נמצאו אפילו 35 שנים אחרי נטישת הכרם. זרעי החרוב מופצים על-ידי יונקים כמו חזירי בר ובקר האוכלים את הפירות הבשלים לאחר שנשרו לקרקע ואולי גם על-ידי עטלפים אוכלי פירות. בלוטי האלון מופצים על-ידי עורבנים (van der Pijl, 1982) האוספים אותם וטומנים אותם בקרקע. נראה שהעורבנים לא נמשכו לכרם מסיבה כל שהיא משום שבניגוד לכך בכרם זיתים ובמטע שזיפים סמוכים נמצאו נבטים רבים של אלונים. האם יחדרו האלונים, הנחשבים למרכיב עיקרי במיני הקיימא, לחלקות בעתיד? זו שאלה מעניינת שהתשובה לה לא תוכל היבדק באתר המחקר משום שלאחר השלמת המחקר הוא הפך לשדה מרעה.
ספרות:
זהרי מ 1959 גיאובוטניקה (מהדורה שניה) ספרית פועלים, תל אביב.
חריף י 1974 התפתחות מרכיבי הגריגה והחרש הראשיים: בשנתם הראשונה וחשיבותה בקביעת הטורים הסוקצסיונים בהרי יהודה . תיזה לדוקטורט, האוניברסיטה העברית ירושלים.
יצחקי ע 1986הפצת זרעים על-ידי ציפורים בחורש הים-תיכוני. תיזה לדוקטורט, האוניברסיטה העברית ירושלים.
פינברון-דותן נ ודנין א 1991 המגדיר לצמחי-בר בארץ-ישראל. כנה. ירושלים.
רבינוביץ-וין א 1986 סלע-קרקע-צומח בגליל. הוצאת הקיבוץ המאוחד ורשות שמורות הטבע, תל-אביב.
———————–
Bazzaz FA 1979 Physiological ecology of plant succession. Annual Review of Ecology and Systematics 10: 351–371.
Billings WD 1938 The structure and development of old field shortleaf pine stands and certain associated physical properties of the soil. Ecol. Monogr. 8: 437-499.
Buell MF Buell HF Small JA and Siccama TG 1971 Invasion of trees in secondary successiorr on the New Jersey Pedimont. Bull. Torr. Bot. Club 98: 67-74.
Chapin FS Walker LR Fastie CL and Sharmen LC 1994 Mechanisms of primary succession following deglaciation at Glacier Bay, Alaska. Ecol. Monogr. 64: 149-175.
Clements FE 1904 The development and structure of vegetation. Botanical Survey of Nebraska 7. Studies in the Vegetation of the States. Lincoln, NE.
Clements FE 1916 Plant succession.Carnegie Institute Washington Publications, Washington, DC.
Connell JH and Slatyer RO 1977 Mechanisms of succession in natural community stability and organization. Am. Nat. 111: 1119-1144.
Debussche M and Isenmann P 1994 Bird-dispersed seed rain and seedling establishment in patchy Mediterranean vegetation. Oikos 69: 414-426.
Debussche M and Lepart L 1992 Establishment of woody plants in Mediterranean old fields: Opportunity in space and time. Landscape Ecol, 6: 133-145.
Debussche M Escarre J and Lepart J 1982 Ornithochry and plant succession in Mediterranean abandoned orchards. Vegetatio 48: 255-266.
Debussche M Lepart L and Molina J 1985 La dissemination des plantes a fruits charnus par les oiseaux: role de la structure de la vegetation et impact sur la succession en region mediterraneenne. Acta Oecol.IOecol. Gener 6: 6580.
Drury WH and Nisbet ICT 1973 Succession. J. Arnold Arbor. 54: 331-368.
Egler FE 1954 Vegetation science concept. I. Initial floristic composition – A factor in old field vegetation development. Vegetatio 4: 412-417.
Escarre J Houssard C and Debussche M 1983 Evolution de la vegetation et du sol apres abandon cultural en region mediterraneenne: etude de succession dans le Garrigues du Montpellierais (France). Acta Oecol. 4: 221-239.
Gill DS and Marks PL 1991 Tree and shrub seedling colonization of old fields in central New York. Ecol. Monogr. 61: 183-205.
Glenn-Lewin DC and van der Maarel E 1992 Patterns and processes of vegetation dynamics. In: Glenn-Lewin D C Peet R K & Veblen T T (eds.) Plant succession theory and prediction,pp. 11-59. Chapman & Hall, London.
Glenn-Lewin DC Peet RK and Veblen TT 1992 Plant succession theory and prediction. Chapman & Hall, London.
Guevara S Meave J Moreno-Casasola P and Laborde J 1992 Floristic composition and structure of vegetation under isolated trees in neotropical pastures. J. Veg. Sci. 3: 655-664.
Guevara S Purata S and van der Maarel E 1986 The role of remnant trees in tropical secondary succession. Vegetatio 66: 74-84.
Holl D 1998 Do bird perching structures elevate seed rain and seedling establishment in abandoned tropical pasture?. Restoration Ecology, 6: 253-261.
Holl KD Loik ME Lin EH and Samuels IA 2000 Tropical montane forest restoration in Costa Rica: overcoming barriers to dispersal and establishment. Restoration ecology, 8: 339-349.
Holthuijzen AMA and Sharik IL 1985 The avian seed dispersal system of eastern red cedar (Juniperus virginiana). Can. J. Bot. 63: 1508-1515.
Horn HH 1981 Succession. In: May, R.M. (ed.) Theoretical ecology principles and applications. Blackwell, Oxford.
Houssard C Escarre J and Romane F 1980 Development of species diversity in some Mediterranean plant communities. Vegetatio 43: 59-72.
McClanahan TR and Wolfe RW 1993 Accelerating forest succession in a fragmented landscape: The role of birds and perches. Conserv. Biol. 7: 279-288.
McCook LJ 1994 Understanding ecological community succession: Causal models and theories, a review. Vegetatio 110: 115-147.
McDonnell MJ and Stiles EW 1983 The structural complexity of old field vegetation and the recruitment of bird- dispersed plant species. Oecologia 56: 109-116.
Myster RW and Pickett STA 1994 A comparison of rate of succession of over 18 yr in 10 contrasting old fields. Ecology 75: 387-392.
Noble IR and Slatyer RO 1980 The use of vital attributes to predict successional changes in plant communities subject to recurrent disturbances. Vegetatio 43: 5-21.
Odum EP 1969 The strategy of ecosystem development. Science 164: 262-270.
Oosting HJ 1942 An ecological analysis of the plant communities of Pedimont, North Carolina. Am. Midl. Nat. 28: 1-126.
Pausas JG Bladé C Valdecantos A Seva JP et al. 2004 Pines and oaks in the restoration of Mediterranean landscapes of Spain: new perspectives for an old practice – a review. Plant Ecology, 171: 209-220.
Pickett STA 1976 Succession: An evolutionary interpretation. Am. Nat. 110: 107-119.
Pickett STA 1982 Population patterns through twenty years of old field succession. Vegetatio 49: 45-59.
Pickett STA 1989 Space-for-time substitution as an alternative to long term studies. In: Likens G E (ed.) Longterm studies in ecology, pp. 110-1335. Springer-Verlag, New York, NY.
Tatoni T and Roche P 1994 Comparison of old-field and forest revegetation dynamics in Provence. J. Veg. Sci. 5: 295-302.
Tilman D 1987 Secondary succession and the pattern of plant dominance along experimental nitrogen gradients. Ecol. Monogr. 57: 189-214.
Tilman D 1988 Plant strategies and the dynamics and structure of plant communities. Monographs in Population Biology 28, Princeton Univ. Press, Princeton, NJ.
van Andel J Bakker JP and Grootjans AP 1993 Mechanisms of vegetation succession: A review of concepts and perspectives. Acta Bat. Neerl. 42: 413-433.
van der Pijl L 1972 Principles of dispersal in higher plants. Springer-Verlag, Berlin.
תודות
תודותינו נתונות ליפעת כהן על עבודת השדה ול M. Deboussche על הערותיו על כתב היד המקורי.
================
נספח 1. רשימת מיני הצמחים הרב שנתיים שנמצאו בכרם הנטוש, צורת-החיים של כל מין, דרך הפצת הזרעים שלו ורשימת מיני הציפורים שנצפו ויכולות לשמש מפיצות של הזרעים (על פי יצחקי 1986). קיצורי שמות הציפורים: אדום חזה (אח), בולבול ממושקף (במ) חכלילית סלעים (חס), חכלילית עצים (חע), קיכלי רונן (כר), סבכי אפור (סאפ), סבכי החורש (סח), סבכי הטוחנים (סט), סבכי שחור-כיפה (סשכ), סבכי שחור-ראש (סשר), סלעית אירופית (סאר), שחרור (שח), סלעית אירופית (סא).
| מין הצמח | צורת החיים | הפצת זרעים | ציפורים מפיצות |
| אספרג החורש | מטפס | ציפורים | אח, במ, סשר |
| קידה שעירה | שיח | עצמית | |
| חרוב מצוי | עץ | יונקים | |
| רקפת מצויה | גיאופיט | עצמית | |
| זקנן שעיר | עשבוני רב שנתי | רוח | |
| טיון דביק | בן שיח | רוח | |
| צמרנית הסלעים | בן שיח | רוח | |
| יערה איטלקית | מטפס | ציפורים | סשר |
| שבטן לבן | בן שיח | ציפורים | חע, כר, סשכ, שח |
| אלת המסטיק | שיח | ציפורים | אח, במ, כע, כס, כר, סשכ, סשר, שח |
| אלה ארץ-ישראלית | עץ | ציפורים | חע, סאפ, סח, סט, סשכ, סשר |
| פרסיון גדול | מטפס | ציפורים | לא ידוע |
| אשחר רחב עלים | עץ | ציפורים | במ, סשכ, סשר, שח |
| אשחר ארץ-ישראלי | שיח | ציפורים | אח, במ, כס, כע, כר,סאפ, סח, סט, סשכ, סשר, שח |
| פואה מצויה | מטפס | ציפורים | אח, במ, סח, סשכ, סשר, שח |
| סירה קוצנית | בן שיח | עצמי | |
| קיסוסית קוצנית | מטפס | ציפורים | במ, כר, סשכ, שח |
| אחירותם החורש | שיח | עצמי | |
| טמוס מזרחי | מטפס | ציפורים | לא ידוע |
==================
כל הזכויות שמורות ל"כלנית" ©
ציטוט: נאמן ג ויצחקי ע 2016 התנחלות צמחי בר, סוקצסיה משנית, בכרם גלילי נטוש. כתב-עת "כלנית" מספר 3.
https://www.kalanit.org.il/?p=7891&preview=true
==================










