עודד כהן, המכון לחקר הגולן, אוניברסיטת חיפה, קצרין. המחלקה ללימודי ארץ ישראל, מכללת כנרת. Odedic@gmail.com
יוסי ריוב, המכון למדעי הצמח וגנטיקה בחקלאות ע"ש רוברט ה. סמית, הפקולטה לחקלאות, מזון וסביבה ע"ש רוברט ה. סמית, האוניברסיטה העברית בירושלים, רחובות. riov@agri.huji.ac.il
תקציר: במאמר זה מובאת סקירה של ממצאים שדווחו בספרות המדעית וממצאים של סקרי שדה לקביעת המעמד האקולוגי של אקליפטוס המקור בישראל. אקליפטוס המקור ניטע בארצות רבות בעולם ובהיקפים גדולים. אף על פי כן, הדיווחים על פלישה של מין זה בעולם, במובן של התפשטות בקנה מידה רחב הם מעטים מאוד, ובוודאי ביחס להיקף הנטיעות העולמי. מצב דומה קיים בישראל. אוכלוסיות צפופות של עצים שמקורן בריבוי טבעי (זריעים) המרוחקים מצמחי אם נצפו עד כה אך ורק בנחל תבור. הנתונים בספרות וכמו גם סקרי השדה, תומכים בכך שאקליפטוס המקור ראוי למעמד של מין שהתאזרח בישראל ולא למעמד של מין פולש.
================
תקציר
אקליפטוס המקור (Eucalyptus camaldulensis), העץ שנודע "מצמרתו ומעלה", הוגדר לאחרונה כמין פולש, הגדרה שעשויה לגרום לנגזרות יישומיות בארץ ובעולם. הגורמים להגדרתו של המין כפולש כוללים התייחסות לשטח תפוצתו הטבעי, דיווחים על פלישה שלו בארץ ובעולם, והתייחסות להשלכותיו על המערכות האקולוגיות. סקר ספרות עדכני מלמדים שאף על פי שהמין ניטע במספר רב של ארצות ובהיקפים גדולים, הפלישה שלו בעולם היא חריגה. התפשטות של אקליפטוס המקור בהיקף נרחב קיימת אך ורק בדרום אפריקה ובמידה מסוימת לאורך החוף הדרומי של חצי האי האיברי ובקליפורניה. מקומות אלה מתאפיינים בפיזור משקעים במשך רוב חודשי השנה, שלא כמו בישראל, שבה העונה היבשה מתפרשת על פני חמישה חודשים במהלך השנה. במהלך החורף של 2016 ערכנו סקר שדה במספר רב של אתרים בישראל, שבהם דווח על פלישה של אקליפטוס המקור: שמורת תל-דן, גן לאומי הרי יהודה, גדות נחל כסלון, שמורת לימן, שמורת כרי נעמן, שמורת השיטה המלבינה באשדוד, נחל שורק, נחל יבנאל, נחל דליות ונחל משושים. מהממצאים עולה, שקיים פער ניכר בין הדיווחים על פלישה למצב בפועל. התפשטות מסוימת של אקליפטוס המקור נראתה באזור נחל דליות, שם נמצאו עשרות זריעים במרחק של עד כמה מאות מטרים מהחורשות הוותיקות, שניטעו עוד בתקופה הסורית. חורשות צפופות במרחק הגדול מ- 30 מ' מצמחי האם לא נצפו כלל בסקר. למעשה הדיווח היחיד על ריבוי טבעי של אוכלוסיה צפופה במרחק מצמחי האם ידוע בנחל תבור בו טופלו העצים בהצלחה אך בעלות גבוהה. לעומת זאת, קיימת תופעה של ריבוי טבעי של עצי אקליפטוס בתוך גבולות החורשות הנטועות, בין חופות העצים ובסמוך להם, בעיקר לצידי כבישים ובמורדות נחלים לאורך פשטי ההצפה שלהם. תופעה זו חוזרת על עצמה הן באגן הניקוז המערבי (בנחלים נעמן, קישון, ירקון ושורק), והן באגן הניקוז המזרחי (בנחלים דליות ותבור). בדומה לממשק האקטיבי הנחוץ לדילול החורש הים תיכוני, ממשק אקטיבי נחוץ גם לטיפול בזריעים במקומות רגישים להתפשטותם. פעולות לוויסות של האוכלוסיה הן מפתח לפתרון המחלוקת בין שימור המגוון הביולוגי מחד, לבין שימור ערכי המורשת ושירותי המערכת האקולוגית של אקליפטוס המקור מאידך.
נטיעות אקליפטוס המקור ומעמדו בישראל – סלע המחלוקת
אקליפטוס המקור (Eucalyptus camaldulensis) הוא עץ ממשפחת ההדסיים שמוצאו מאוסטרליה, ומשמש בעיקר לצל, לנופש, למרעה דבורים, לנטיעות כעץ שדרה וליעור. אקליפטוס המקור הובא לארץ כבר בשנות השמונים של המאה ה- 19, בהנחה שהוא יכול לשמש אמצעי לייבוש ביצות (רוזן 1991). בהמשך ניטעו בארץ מינים שונים של הסוג אקליפטוס בהיקפים גדולים על ידי קק"ל (הקרן הקיימת לישראל) וממשלת המנדט הבריטי. מלבד תועלותיו לשירותי המערכת האקולוגית, אקליפטוס המקור אדיר הממדים, הפך גם לסמל מיתי של המפעל הציוני ויישוב הארץ. לעץ זה שמור אף מקום של כבוד בסיפור חייו של המרגל הנודע אלי כהן, שכפי שמספרת האגדה, נעזר בחורשות אקליפטוסים ל"סימון" בסיסי הצבא הסורי. מנתוני הנטיעות של הקק"ל מהתקופה שלפני קום המדינה ועד לימים אלו, עולה ששטחי הנטיעות של אקליפטוס המקור ועומדים בהם תצורת הצומח היא "אקליפטוסים" (לרוב אקליפטוס המקור ומיני אקליפטוס נוספים) מסתכמת ב- 77,844 דונם. החל משנות ה- 80 של המאה הקודמת, אקליפטוס המקור ניטע ליער משקי (חקלאי) לתפוקת עץ במישור החוף ובעמקים הפנימיים, אולם נטיעות אלו פסקו בשנים האחרונות בשל העדר כדאיות כלכלית. נכון להיום אקליפטוס המקור משמש בארץ בהיקף קטן יחסית לייעור. בין השנים 2011 ל- 2016 ניטעו 3,753 דונם של חורשות המבוססות על מיני אקליפטוסים בלבד, 81% מהן בדרום הארץ בהן נטעו מינים העמידים ליובש.
לאחרונה הועלה חשש שהעץ מזריע את עצמו ומתפשט בעיקר לאורך נחלים ובבתי גידול לחים. בשל הדיווחים על התפשטותו ולאור תכונותיו האללופתיות, העץ סווג בישראל כמין פולש, יחד עם רשימת מיני צמחים פולשים הנחשבים כעשבים רעים, המאיימים על שמירת הטבע (environmental weeds) ועל החקלאות (agricultural weeds) (דופור-דרור, 2010). באותה רשימת מיני צמחים פולשים, נכללים לדוגמה טיונית החולות (Heterotheca subaxillaris) המאיימת על שימור בתי הגידול החוליים, אמברוסיה מכונסת (Ambrosia confertiflora) הנחשבת למפגע סביבתי וחקלאי גם יחד, חסת המים (Pistia stratiotes) הנחשבת לאחד מבין העשבים הרעים הסביבתיים הקשים בעולם, ושיטה כחלחלה (Acacia saligna), שמהווה עשב רע סביבתי ביערות, בשטחים פתוחים ולאורך צידי תשתיות. מאמר זה דן באקליפטוס המקור מבחינת הגדרתו כמין פולש והערכת האיום שלו על המערכות האקולוגיות בישראל.
הקריטריונים להגדרת האקליפטוס כמין פולש בישראל – המצב הקיים
אקליפטוס המקור הוגדר כ"מין פולש" (invasive) ברשימה שפורסמה על-ידי רשות הטבע והגנים (רט"ג) בספר "צמחים פולשים בישראל" (דופור-דרור ,2010, Dufour-Dror, 2012) . מין פולש הוגדר כ "מין של צמח שהוא גם זר לסביבה שבה הוא חי וגם הרסני בצורה כלשהי לסביבה ולאורגניזמים המקומיים" (Wilson, 2003 בתוך דופור-דרור, 2010). הגדרה זו זהה גם במהדורה המחודשת באנגלית (Doufor-Dror, 2012). רמת האיום של אקליפטוס המקור הוגדרה "רמה 3", שמשמעותה "צמח הפולש בשטחים טבעיים ובהם הוא יוצר עומדים צפופים אך אינו בעל תכונות אללופתיות, מציב איום אקולוגי גבוה יותר – רמה 3. הסיבה לכך היא שדחיקה מגיעה לשיאה כאשר צמח פולש יוצר עומדים צפופים בעלי כיסוי סגור". כיוון שהאקליפטוס ידוע ביכולותיו האללופתיות, אנו מניחים שכוונת הסוקר הייתה לדרג את האקליפטוס ברמת סיכון 4, המייחסת למינים גם תכונה של יכולות אללופתיות. הקריטריונים להערכת האיום של מינים פולשים מתוארים בספר. להלן התייחסותו לאקליפטוס המקור:
- תפוצתו הטבעית של אקליפטוס המקור באוסטרליה מצביעה על סתגלנות רבה. תפוצתו הטבעית מתפרשת על פני מגוון רחב של אזורים אקלימיים, כמעט בכל היבשת.
- העץ מסוגל לצמוח במגוון בתי גידול באקלים הים תיכוני ובאקלים הצחיח למחצה בישראל.
- השפעות הצמח על הסביבה: העץ ניחן בתכונות אללופתיות, ולכן עולה החשש לפגיעה במגוון המינים. כמו כן, אפשר שהחורשות הצפופות מביאות לירידת מפלס המים ולייבוש חלקי של בתי גידול.
- העץ מוגדר כפולש גם במקומות אחרים בעולם כדוגמת קליפורניה, הוואי ודרום אפריקה, בה אף נאסרה שתילתו ליד מקורות מים.
- העץ פולש לשטחים טבעיים בישראל, דוגמת נחל שורק, שמורת כרי נעמן ושמורת תל-דן.
פוטנציאל הפלשנות ורגישות לפלישה של אקליפטוס המקור
מיני אקליפטוס שונים ניטעו בהיקף נרחב בכל העולם. נכון לשנת 2008, סך השטח הכללי של נטיעות אקליפטוס מחוץ לאוסטרליה הסתכם ב- 200 מיליון דונם (Booth, 2014). לכאורה, בשל גודל הקבוצה הטקסונומית (מעל ל-800 מינים באוסטרליה ובסביבתה) וגם בשל היקף הנטיעות העולמי, אפשר היה לצפות, שלסוג אקליפטוס תהיה נציגות רחבה בפלישה הכוללת של עצים ושיחים בעולם. אולם, היקף הפלישה של מיני האקליפטוס השונים נמוך יחסית בהשוואה לסוגים אחרים כמו ינבוט (Prosopis) ושיטה Acacia (לפי Rejmánek and Richardson, 2011; Booth, 2012). הספרות המדעית מבדילה בין פוטנציאל הפלשנות (invasiveness), המתייחס ליכולת המינים הזרים להתקדם בשלבי הפלישה הביולוגית על סמך תכונותיהם, לבין פוטנציאל הנפלשות (invasibility), המתייחס לרגישות בית הגידול לפלישת מינים זרים (Alpert et al., 2010). ריימנק וריצרדסון (2011 ,Rejmanek and Richradson) מסבירים את פוטנציאל הפלישה הנמוך של מיני האקליפטוס ביכולת מוגבלת של הפצה לטווח רחוק, שרידות נמוכה של הנבטים והעדר אקטומיקוריזה מתאימה בבתי הגידול החדשים. להלן התייחסות מפורטת למספר הגורמים המקשים על הפלישה של אקליפטוס המקור בסביבות חדשות:
- מרחק הפצה מוגבל – היכולת של מינים פולשים לבסס אוכלוסיות במספר רב של מוקדים תוך פרק זמן קצר יחסית, קשורה במידה רבה ביכולתם להפיץ זרעים ולהתנחל במרחק רב מצמח האם (Lake et al. 2004). מינים המותאמים להפצה לטווח רחוק מייצרים בדרך כלל כמויות גדולות של זרעים קטנים המצוידים באמצעי תעופה להפצה על ידי הרוח, כמו לדוגמה כנפון בזרעים של כנפון זהוב (Verbesina encelioides) או ציצית בזרעים של טיונית החולות. בשטחים שבהם עושר המשאבים גדול, ולכן כיסוי הצומח גדול, כמו לדוגמה לאורך נחלים ובתי גידול לחים, ישנה העדפה להפצה לטווח רחוק על ידי פירות בשרניים המותאמים להפצה על ידי יונקים או זרעים בעלי אריל (aril) שומני אדום וגדול, שהוא אטרקטיבי למגוון גדול של ציפורים (Lake et al. 2004). רוב מיני השיטה האוסטרליים הידועים כפולשים בעולם, הם בעלי אריל המותאם להפצה על-ידי ציפורים, אם כי קיימת גם הפצה על ידי נמלים ועל-ידי וקטורים נוספים. בנוסף לכך, צמחים פולשים בבתי גידול לחים מתפשטים לטווח קרוב באמצעות סורי שורש ולטווח רחוק באמצעות יונקים המפיצים את פירותיהם הקוצניים, וכן על ידי חלקי צמח הניתקים ומוסעים עם זרמי המים, כמו לדוגמה אמברוסיה מכונסת (דופור-דרור 2015). הזרעים של מרבית מיני האקליפטוס, ובכלל זה אקליפטוס המקור, הם קטנים מאוד יחסית למינים אחרים וחסרי התאמות להפצה למרחק, כמו כנפיים להפצה על ידי הרוח או רקמות בשרניות להפצה על ידי בעלי חיים. זרעי אקליפטוס נאספים אמנם על-ידי נמלים, אולם לא נצפתה נביטה שלהם לאחר צבירתם בסביבת הקינים (O’Dowd and Gill, 1984). מקובל אמנם שזרעים קטנים מופצים על ידי הרוח למרחקים גדולים יותר מאשר זרעים גדולים, אך עם זאת לעתים ההבדל בטווח ההפצה אינו גדול (Ladd and Cappuccino, 2005 וספרות המצוטטת שם). מרבית המחקרים מעידים על כך שטווח ההפצה האפקטיבי של זרעי מיני אקליפטוס שונים הינו קטן יחסית (Rejmanek and Richardson, 2011). למשל, עפ"י (Cremer, 1977), במינים רבים של אקליפטוס, מרחק זה הינו פחות מפעמיים גובה עצי האם. ממצאים אלו תואמים לממצאי מחקרים על מהירות הנפילה המקסימלית של זרעים לקרקע. מהירות נפילה גבוהה יחסית, משמעה מרחק הפצה קטן עלידי הרוח. מהירות הנפילה המקסימלית של רוב הזרעים של מיני האקליפטוס היא בטווח של 5-2 מ' לשנייה, בעוד שבאורנים לדוגמה המהירות היא 1.5-0.7 מ' לשנייה (Rejmánek and Richardson, 2011). היותם של זרעי אקליפטוס המקור קטנים מאד באופן יחסי, משפיעה גם על שרידותם כנבטים. מזרעים קטנים מתפתחים לעתים קרובות נבטים קטנים, שכושר התחרות והעמידות שלהם קטנים יחסית (Moles and Westoby, 2004). מלבד גורמי ההפצה הטבעיים, האדם ממלא אף הוא תפקיד חשוב בהפצה של צמחים בשלבי הגעתם והתבססותם בבתי גידול חדשים (Von Der Lippe and Kowarik, 2007). כיוון שהזרעים הזעירים של אקליפטוס חלקים ואינם מצוידים בקוצים, האדם וכמוהו גם בעלי החיים, אינם משמשים וקטורים יעילים להפצתם. אמנם ישנה אפשרות שזרעים יופצו באמצעות זרמי מים למרחק רב, וכן בעזרת בוץ הנדבק לפרסות בעלי החיים, לסוליות הנעליים של מטיילים ולדפנות צמיגי רכבים, אולם כפי שיוסבר להלן, פוטנציאל ההישרדות של הזרעים והנבטים נמוך, והכמות של הזרעים המגיעים למוקד מרוחק מצמחי האם קטנה מדי כפי הנראה לצרכי התנחלות. למרות שעץ בודד של אקליפטוס המקור מייצר מאות אלפים ואף מעל מיליון זרעים (א. שמידע, מידע בע"פ), בישראל נראים בשטח רק נבטים מועטים. מכאן שאקליפטוס המקור מראה הצלחה מועטה בהתנחלות במרחק גדול מצמחי האם. בנוסף למגבלות של תפוצה לטווח רחוק שתוארו לעיל, ישנם גורמים נוספים המשפיעים על יכולת הפלישה של אקליפטוס המקור: זרעי אקליפטוס המקור הם זעירים, בעלי רקמת הזנה (endosperm) שאינה מפותחת ולכן לא תומכת בהצצה מעומק הקרקע. דווח שבתחום תפוצתו הטבעי מרבית הזרעים נותרים על האדמה חשופים לגרניבוריה (טריפת זרעים), לפגיעה על-ידי פתוגנים ולקטסטרופות אפשריות כמו שריפה (Potts and Wiltshire, 1997). המספר הזעום של הנבטים שנצפו בארץ במרחק מצמח האם, מרמז על כך שהפגיעה בזרעים אינה ייחודית לתחום התפוצה הטבעי ומתרחשת גם בישראל ובמקומות אחרים בעולם. הצלחת ההתנחלות של אקליפטוס המקור במרחק גדול מצמח האם באזור התפוצה הטבעי שלו, חלה רק במקרים נדירים, כאשר כמות גדולה של זרעים מופצת על-ידי זרמי מים לסביבה חשופה, שלרוב מתהווה מיד לאחר שריפה (Potts and Wiltshire, 1997). עם זאת, שילוב של קרקע חמה ולחה ידוע כגורם המקצר את אורך החיים של זרעים במספר רב של מינים (Chambers and Macmahon, 1994), לרבות אקליפטוס המקור (Roberts and Marston, 2000), כפי הנראה כתוצאה מהגברת נביטה, פגיעה הידרותרמית ותקיפת פתוגנים.
- שרידות נמוכה של נבטים – בדומה לשרידות הזרעים, גם שרידות הנבטים תלוייה בתנאי הסביבה. תחרות עם צומח צפוף, בעיקר על משאבי אור ומים, מגבילה מאד את שרידות הנבטים. בנוסף, בהעדר רקמת אנדוספרם מפותחת, בקרקע לחה הנבט חייב לבצע הטמעה לאחר ההצצה לשם שרידותו. לכן, נביטה בקרקע לחה תחת צל כבד האופייני לצומח צפוף, משולה אף היא לנביטת התאבדות בשל רמת הטמעה נמוכה. קרקע לחה מסייעת לנבטים של אקליפטוס להעמיק שורש במהירות, ולכן תנאים בהם הקרקע לחה וחשופה לקרינת השמש, עשויים להיות רגישים יותר להתנחלות של אקליפטוס (Calviño-Cancela and Rubido-Bará 2013). באזורים בהם הקיץ יבש, כמו למשל באזורי אקלים ים תיכוני, הסיכוי לשרידות הנבטים של אקליפטוס המקור קטן ביותר, גם בהעדר צומח טבעי מתחרה. שולי תעלות וגדות נחלים באקלים הים תיכוני של ישראל מכוסים לרוב בצומח סבוך של צמחי גדות. אחד המאפיינים של צמחיית בתי גידול לחים הוא, שהצמחים מתרבים גם וגטטיבית ויוצרים עומדים צפופים המחזקים את העמידות הביוטית של בית הגידול בפני פלישה של צמחים זרים, ובפרט כאשר מדובר במינים בעלי זרעים קטנים. על אף זאת, מכלול של גורמים הקשורים במשטר ההפרעות הגבוה בבתי גידול לחים, כמו שינויים במשטר חומרי הזנה, זמינות מים גבוהה ושינויים במשטר הזרימה של המים המשמשים כווקטור להפצה, מגדילים את הרגישות של גדות הנחלים לפלישה של צמחים זרים (Zedler and Kercher, 2004). סילוק הצומח הטבעי פוגע בחסינות הביוטית של בית הגידול לפלישה ופותח חלון הזדמנויות לפלישה של צמחים זרים, לרבות צמחים המייצרים כמויות גדולות של זרעים קטנים שאינם בעלי תרדמה, אך מצטיינים בהפצה לטווח רחוק כמו מיני אשל (Tamarix) ואסתר מרצעני (Aster ageratoides). משתמע מכך, שיעילות ההפצה לטווח רחוק, שהיא חסרה באקליפטוס המקור, הינה גורם המגדיל את ההסתברות להצלחת פלישה של צמחים בעלי זרעים זעירים בבתי גידול לחים. מלבד התחרות עם מיני צומח מקומי, הרביבוריה יכולה אף היא למלא תפקיד בחסינות הביוטית של בית גידול לפלישה של של מיני אקליפטוס. רעיה נמצאה כגורם המשפיע במובהק על קטילה של נבטי אקליפטוס גלובולוס בבתי גידול לחים וחשופים באזור הים תיכוני של צ'ילה (Becerra and Bustamante, 2008), אולם השפעתה של הרעיה על נבטי אקליפטוס המקור בישראל אינה ידועה.
- העדר אקטומיקוריזה מתאימה – הסוג אקליפטוס מקיים יחסי גומלין עם מיני אקטומיקוריזה שמקורם באוסטרליה. אף על פי שניתן לגדל במשתלות אקליפטוסים ללא אקטומיקוריזה, ישנה דעה, שהסימביוזה עם אקטומיקוריזה חיונית להתנחלות ולהתבססות של הנבטים בתנאים טבעיים. העדר אקטומיקוריזה מתאימה מחוץ לגבולות הנטיעה תורמת לכך שנבטי האקליפטוס מתקשים לשרוד ולהתבגר במרחק מצמח האם (Rejmánek and Richardson, 2011). חוקרים מספרד העלו את החשש, שהתקדמות הפלישה של מיני אקטומיקוריזה אוסטרליים בסביבה חדשה, עלולה להאיץ את ההתנחלות של מיני אקליפטוס (Díez, 2005). עם זאת, (Rejmánek and Richardson (2011 מסייגים ומציינים, שהתנחלות אקליפטוס מתחילה לרוב בקשרים עם פטריות אנדומיקוריזה לא ספציפיות, ולכן לדעתנו יש לייחס לנוכחות האקטומיקוריזה השפעה משנית בהשוואה לגורם האקלים, כפי שיוסבר בהמשך.
- העדר אללופתיה אפקטיבית – בספרות מופיעים מספר דיווחים על האללופתיה של אקליפטוס המקור (Ruwanza et al., 2015 וספרות המצוטטת שם), שיש המייחסים לה חשיבות לגבי כושר הפלישה של צמחים זרים, מאחר והיא מקטינה או מונעת את התחרות עם הצומח הטבעי (Callaway and Aschehoug, 2000). מרבית המחקרים בנושא זה, בדומה לזה של Ruwanza וחוב' (2015), שעסקו בהערכת הפוטנציאל האללופתי של אקליפטוס המקור על ידי בחינה של ההשפעה של מיצויי עלים על צמחי בוחן בקרקעות שונות ובעוצמות שטיפה שונות, לא בוצעו בשדה אלא בתנאים המדמים תנאי שטח. בעבודות אלו נמצא, שמיצויים אלו עיכבו את הנביטה ו/או את התפתחות של צמחי הבוחן. תוצאות דומות התקבלו גם במחקרים שנערכו בארץ, אולם ההשפעה של מיצויי העלים של אקליפטוס המקור הייתה בדרך כלל קטנה מזו של מיצויי עלים של מינים אחרים של אקליפטוס ורסק עלים של מין זה שנטמן בקרקע לא מנע התפתחות עשבים רעים בגידולים חקלאיים (רגיל, 2002, גליקסמן ,2007). למיטב ידיעתנו, הדיווח המדעי היחיד בו מצויינת השפעה אללופתית של אקליפטוס המקור בשטח מתייחס לנטיעות של מין זה שנעשו בקליפורניה (del Moral and Muller, 1970). ההשפעה האללופתית נצפתה בקרקעות כבדות באזורים בהם רמת המשקעים נמוכה יחסית, תנאים שאפשרו הצטברות חומרים אללופתיים בשכבת הקרקע העליונה בגלל העדר שטיפה. תופעת האללופתיה לא נצפתה באזורים סמוכים בהם היו קרקעות קלות יחסית. בתצפיות שנערכו על-ידינו באזורים שונים בארץ, לא נמצאו תופעות של העדר צמחיה מתחת לחופה של אקליפטוס המקור, שניתן ליחס אותן בוודאות להשפעה אללופתית. תופעה זו נצפתה על-ידינו גם באזורים שחונים יחסית. שבהם קיימת אללופתיה של מיני אקליפטוס אחרים. מקובל שתנאי יובש מגבירים את ההשפעה האללופתית של צמחים בשל העדר שטיפה של חומרים אללופתיים (May and Ash, 1990), וכן בשל ייצור מוגבר שלהם על-ידי הצמח בתגובה לעקת יובש (Pedrol et al., 2006).
מן הראוי לציין, שהערכת סיכון נמוכה אין משמעותה שפלישת מיני אקליפטוס אינה אפשרית, שכן ההערכה כשמה כן היא, כלומר היא מבוססת על הסתברות. עם זאת, לגבי אקליפטוס המקור הערכת הסיכון הנמוכה באה לידי ביטוי גם בשטח, מאחר שלמרות היקף הנטיעות העצום של מין זה ונוכח המספר הרב של הזרעים שהוא מייצר, הפלישה שלו התממשה רק במקרים בודדים בשל תנאי סביבה מיוחדים.
אקליפטוס המקור כמין פולש בעולם – היבטים אקולוגיים וגיאוגרפיים
אקליפטוס המקור הוא אחד מבין מיני האקליפטוס הדומיננטיים שניטעו מחוץ לאוסטרליה (Booth, 2014). להלן מובאים דיווחים על הפלישה של אקליפטוס המקור בארצות שונות בעולם. יש לציין, שלעיתים ישנה אי התאמה בדיווחים על הפלישה של מין זה בארצות מסוימות, שנובעת מהעדר נתונים מבוססים על מידת הפלישה, מאבחנה לא מספקת בין התאזרחות לבין פלישה ומהתייחסות להשפעות של הפלישה ולא להיקפה. סקירה עולמית של CABI) Centre for Agriculture and Biosciences International) לגבי נטיעות ופלישה של אקליפטוס המקור בעולם מציינת את הממצאים הבאים: המין ניטע ב- 115 ארצות מחוץ לאוסטרליה וסביבתה, מתוכן 54 ארצות באסיה, 38 ארצות באפריקה, שש ארצות בצפון אמריקה, תשע ארצות בדרום אמריקה ושמונה ארצות באירופה. מתוך 115 ארצות בהן האקליפטוס ניטע כצמח זר (לא כולל אוסטרליה וסביבתה), המין מצוין כפולש בשלוש ארצות בלבד, בבנגלדש, בדרום אפריקה ובקליפורניה. בסקירה שנעשתה בדרום ובדרום-מזרח אסיה (Pallewatta et al., 2003). ארגון המינים הפולשים העולמי (GISP), כלל את אקליפטוס המקור ברשימת המינים הפולשים בפיליפינים, בבנגלדש ובפקיסטן. אנו מצאנו, שכלילת מין זה ברשימת הפולשים בפיליפינים מבוססת בעיקר על השפעתו על בית הגידול, הנובעת מהיות המין וקטור להפצת מזיקים (Joshi, 2006), ולא על בסיס נתונים אודות ההתפשטות שלו בסביבה החדשה. לא הצלחנו למצוא מאמר מדעי מבוסס המאמת את התפשטות אקליפטוס המקור בפיליפינים, בפקיסטן או בבנגלדש. לפי מאגר המידע העולמי של מינים פולשים בעולם (Global Invasive Species Database), ישנם 62 מינים פולשים בבנגלדש, אך אקליפטוס המקור אינו נכלל ביניהם (GISD, 2015).
בניגוד לאמור לעיל, המין מוכר כפולש ללא מחלוקת בדרום אפריקה, שם המין מתפשט בעיקר לאורך נחלים, וההתפשטות מלווה בפגיעה במינים המקומיים (Tererai et al., 2015). יש הסבורים, שמקור הפגיעה קשור ליכולת האללופתית של האקליפטוס (Ruwanza et al., 2015). בכל אופן, כיוון שאקליפטוס המקור בארץ זו מתפשט במוקדים רבים ויוצר אוכלוסיות צפופות במוקדים החדשים (Forsyth et al., 2004), יש בכך כדי לשנות את ההרכב והתפקוד של המערכות האקולוגיות.
לפי אתר DAISIE – Delivering Alien Invasive Species Inventories for Europe) ,DAISIE 2015), אקליפטוס המקור התבסס במספר ארצות באירופה: האיים הקנאריים, יוון, איטליה, פורטוגל, סיציליה, ספרד וטורקיה. אולם המינוח התבסס (established), כפי שנעשה בו שימוש באתר זה, אינו מבדיל בין מינים שהם במעמד של התאזרחות לבין מינים שהם במעמד של פלישה בסביבתם החדשה.
בסקירה של (Rejmánek and Richardson (2011, אקליפטוס המקור מצוין כפולש בקליפורניה, ספרד ודרום אפריקה. אקליפטוס המקור מצוין ברשימת הצמחים הפולשים באנדולסיה ( ana et al., 2005), אך ללא תיאור מפורט של היקף ההתפשטות. נראה, שיותר מאשר היקף ההתפשטות הנוכחי, החוקרים חוששים מפלישה אפשרית בעתיד באזורי החופים הואלבה (Huelva) וקאדיז (Cadiz), ומההשפעות האפשריות של האקליפטוס על הסביבה. החוקרים מציינים, שכריתה והדברה של עצי אקליפטוס בוגרים הינה פעולה מורכבת הכוללת צורך במיכון כבד, והם רואים באקליפטוס המקור פולש בעייתי תוך לקיחה בחשבון של הקושי בטיפול בעצים בוגרים. מלבד רשימת הצמחים הפולשים באנדולסיה (Dana et al., 2005), מצאנו שני מאמרים נוספים, בהם אקליפטוס המקור מאוזכר כפולש בספרד. המאמר הראשון עוסק בהערכת הסיכון לפלישה עתידית של צמחים פולשים, על בסיס ההבדלים בין שטח התפוצה שלהם בהווה לשטח התפוצה הפוטנציאלי (Gassó et al., 2012). אולפ החוקרים אינם מציגים במאמר נתונים עדכניים על מאפייני ההתפשטות של אקליפטוס המקור בספרד. המאמר השני, עוסק בקשר שבין התבססות אקטומיקוריזה שמקורה באוסטרליה לבין התבססות ופלישה אפשרית של מיני אקליפטוס בספרד (Diez, 2005). לגבי המצב כיום החוקרים מדווחים, כי התפשטות עצי אקליפטוס בחצי האי האיברי (ספרד, פורטוגל ואנדורה) מוגבלת לאזורים הסמוכים לחורשות נטועות גדולות. אכן אקליפטוס המקור נמצא ברשימת המינים הזרים שהתאזרחו בפורטוגל (Domingues De Almeida and Freitas, 2006).
בארצות הברית, אקליפטוס המקור נחשד אמנם כפולש בקליפורניה ובפלורידה, אך העדויות לכך הן מעטות מאוד, והדיווחים בספרות תומכים בכך שהמין שם נמצא במצב של התאזרחות ולא במצב של פלישה (Lorentz and Minogue, 2015b). המונח 'התאזרח' מייחס למין תכונת ריבוי עצמאית, כלומר שהמין התגבר על מכשולי הרבייה בבית הגידול, אך לא על מכשולי ההפצה וההישרדות של הפרטים במוקדים החדשים. מין פולש (invasive) לעומת זאת, הינו מין שהתאזרח והתפשט מהמוקדים המבוססים להרבה מוקדים חדשים, שם הוא מעמיד אוכלוסיות המתרבות באופן עצמאי ובצפיפות. לפי הארגון העוסק במינים פולשים בקליפורניה (CAL-IPC) , מין זה מוגדר שם כפולש, שרמת האיום שלו על האוכלוסיות הטבעיות נמוכה (limited). במספר פירסומים מהשנים האחרונות, השתמשו חוקרים במונח מין שהתאזרח (naturalized) כדי לתאר את הסטטוס של אקליפטוס המקור בקליפורניה (Ritter and Yost, 2009, Lorentz and Minogue, 2015a).
מחקר נוסף מייחס רמת סיכון גבוהה לפלישה של מינים שונים של אקליפטוס בפלורידה וביניהם גם אקליפטוס המקור (Gordon et al., 2011). ההערכה נעשתה על סמך ניתוח של 49 מאפיינים, שהוגדרו על בסיס מקורות ספרות ביחס לתכונות האקליפטוס כמו מאפייני בית הגידול בסביבה החדשה, יחסי הגומלין בין השניים ונתונים ביודמוגרפיים הקשורים באזור התפוצה של המין באוסטרליה ובאזור הפלישה בדרום אפריקה (Gordon et al., 2011). לעומת זאת, מחקרים חדשים שהתמקדו בשלבי הנביטה והצימוח של אקליפטוס המקור בשנתו הראשונה הראו, שקיים פער בין מודל ההערכה מבוסס הספרות לבין המצאות נבטים ושרידותם בשטח, המעידים על סבירות נמוכה מאוד לפלישה (Lorentz and Minogue, 2015b). ראשית, מתוך למעלה מ-300 חלקות שנדגמו במגוון בתי גידול בפלורידה לא נמצאו נבטים כלל במרחק מחורשה נטועה. כמו כן נמצא, ששזרוע מכוון של אלפי זרעים הביא לנביטה זעומה בשטח, וששתילים צעירים שניטעו במגוון של נישות אקולוגיות לא שרדו מעבר ל-13 שבועות לאחר הנטיעה. ממצאים אלה עולים בקנה אחד עם ממצאים אחרים המראים, שפוטנציאל הפלישה של מיני אקליפטוס בפלורידה, לרבות אקליפטוס המקור, הוא נמוך מאד (Callaham et al., 2013).
חיפוש במאגרי מידע אקדמיים (SCOPUS, SCIENCE DIRECT, JSTOR) העלה, שישנם מעט מאוד דיווחים על אקליפטוס המקור כפולש (invasive). מרבית המאמרים האלו עוסקים בכלל בפלישה של מזיקים הפוגעים באקליפטוס, כמו לדוגמה הפלישה של צרעת עפצי החטטים בישראל (Protasov et al., 2007). מספר מאמרים עוסקים בהשפעות אקליפטוס המקור על הסביבה, בעיקר בדרום אפריקה, וברוב המאמרים הנושא של מין זה כפולש אינו נושא מרכזי. האמור לעיל מתייחס גם למיני אקליפטוס אחרים. מסתבר שפלישה אקוטית של אקליפטוס המקור, שאינה נתונה במחלוקת, מדווחת אך ורק בדרום אפריקה (Forsyth et al. 2004), ובמידה מסוימת בקליפורניה. יש הסבורים (,Booth 2012), כי הפלישה של מיני אקליפטוס בדרום אפריקה קשורה בהיסטורית הנטיעה שם, שהיא ותיקה ב-50 שנה בהשוואה לאזורים אחרים בעולם (החלה משנות העשרים של המאה ה-19), וכמו גם בהיקף הנטיעות הגדול שם, תוך ציון ש"היקף הנטיעות הגדול ביותר מלווה גם בפלישה הגדולה ביותר." למיטב ידיעתנו, אקליפטוס המקור ניטע בארצות הברית כבר בראשית המאה ה-19 (בדומה לדרום אפריקה), כמו כן הוא ניטע בהיקפים גדולים באזורים רבים בעולם, בעיקר בדרום אמריקה ובאסיה (Turnbull, 1999). לפיכך, מועד הנטיעות והיקפן, לא נראים לנו כגורמים המסבירים כשלעצמם את הפלישה של אקליפטוס המקור בדרום אפריקה.
השוואת התנאים האקלימיים בין ארצות שבהם אקליפטוס המקור התאזרח ופולש, לתנאים בישראל
האקלים ידוע כגורם הראשי המגביל את ההישרדות, הרבייה והיצרנות של צמחים במערכות אקולוגיות יבשתיות. כיוון שאקליפטוס המקור הוא צמח המותאם אבולוציונית לגדות נחלים, גורם המשקעים נראה בעינינו כגורם חשוב ביותר לפוטנציאל ההתנחלות של האקליפטוס בבתי גידול חדשים, ובפרט באזורי אקלים יובשניים למחצה. תפוצת אקליפטוס המקור באוסטרליה היא אכן רחבה מבחינת גודל השטח, אך מוגבלת לרוב לאורך צידי נחלים ושטחים מוצפים. הצפות מהוות את הגורם הקובע את הצלחת ההתבססות של הנבטים באזורים היובשניים והיובשניים למחצה באוסטרליה, כאשר הצפה באביב המוקדם המלווה בקיץ מתון מהווה את התנאים האופטימליים להתבססות נבטים של אקליפטוס (Roberts and Marston, 2011). יערות של אקליפטוס המקור משגשגים בעיקר בדרום מזרח אוסטרליה שם יורדים גשמים במהלך כל השנה, מעל ל- 30 מ"מ בחודש היבש ביותר. ההבדלים בתנאי האקלים בישראל (בית-דגן) בהשוואה לפלורידה (טאמפה), קליפורניה (סאן פרנסיסקו), דרום אפריקה (קייפטאון) וספרד (קאדיז) (איור 1) עשויים להשפיע באופן המשמעותי ביותר על הישרדות נבטי אקליפטוס בתנאים טבעיים. הערים הנבחרות מחוץ לישראל מייצגות אזורים שבהם ידועה התאזרחות או פלישה של אקליפטוס המקור (איור 1) (USDA 2015).
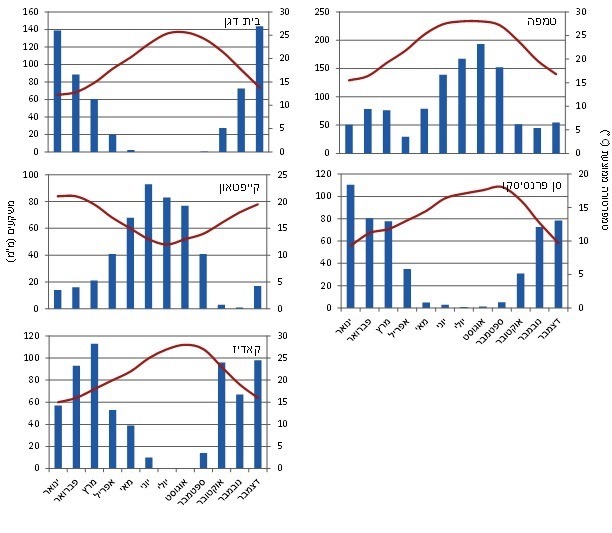
איור 1. נתוני האקלים (טמפרטורה ממוצעת במעלות צלזיוס ומשקעים) במהלך השנה בחמש ערים המייצגות אזורים בהם ידוע שאקליפטוס המקור התאזרח או נחשב למין פולש. הנתונים מתוך אתר ClimaTemps.com (הורדה בתאריך ה- 13.01.2015). הערים: טמפה (Tampa), פלורידה; סן פרנסיסקו (San Francisco), קליפורניה; בית דגן (Bet Dagan), ישראל; קייפטאון (Cape Town) , דרום אפריקה; קאדיז (Cadiz), ספרד.
אזור האקלים הים-תיכוני בישראל (בית-דגן) מאופיין בממוצע משקעים שנתי של 550 מ"מ. הגשמים יורדים אמנם בין ספטמבר למאי, אך הגשמים האפקטיביים (90% מהמשקעים) יורדים במשך חמישה חודשים בלבד, בין נובמבר למרץ. טמפרטורת היממה הממוצעת על בסיס חודשי נמצאת ביחס הפוך לממוצע המשקעים החודשי, בטווח שבין 12 מעלות צלזיוס בינואר ל- 26 מעלות צלזיוס באוגוסט.
האקלים בפלורידה (טאמפה) הוא סוב-טרופי לח עם ממוצע משקעים שנתי של למעלה מ- 1000 מ"מ. הגשמים יורדים במשך כל חודשי השנה. טמפרטורת היממה הממוצעת על בסיס חודשי נמצאת ביחס ישר לממוצע המשקעים החודשי, בטווח שבין 15 מעלות צלזיוס בינואר ל- 28 מעלות צלזיוס באוגוסט. האקלים בפלורידה מזכיר במידה רבה את זה של דרום מזרח אוסטרליה, שם יערות אקליפטוס המקור משגשגים. לאור ההבדלים האקלימיים בין פלורידה לישראל, לא ניתן לדעתנו להקיש מהתבססות אקליפטוס המקור בפלורידה על פוטנציאל הפלישה בישראל.
בהשוואת האקלים הים-תיכוני בין דרום אפריקה (קייפטאון) לישראל (בית-דגן), מסתמנים הבדלים, בעיקר במשך העונה היבשה ובצחיחותה. משך התקופה היבשה בקייפטאון, שבה כמות המשקעים החודשית (המצטברת) קטנה מ-3 מ"מ, מסתכמת בחודשיים בלבד, במהלך החודשים אוקטובר – נובמבר. שני החודשים האלו חלים לאחר חודש שבו יורדים כ-41 מ"מ בממוצע, ערכים שבוודאי עשויים ליצור נגר משמעותי לאורך גדות הנחלים ופשטי הצפה. לעומת זאת, פרק הזמן היבש בישראל מסתכם בחמישה חודשים החל מסוף חודש אפריל, שבו יורדים בממוצע 20 מ"מ באופן בלתי סדיר בהתפלגות רב-שנתית. טמפרטורת היממה המקסימלית הממוצעת על בסיס חודשי במהלך השנה, אינה חופפת את פרק הזמן היבש בקייפטאון, שבו ישנם ערכים של 16-15 מעלות צלזיוס. לעומת זאת, טמפרטורת היממה המקסימלית הממוצעת לחודש במהלך השנה בישראל חופפת את פרק הזמן היבש, שבו מתקבלים ערכים של 26-25 מעלות צלזיוס. מכאן שהעונה היבשה בישראל ארוכה יותר מאשר בקייפטאון וצחיחה יותר. פירושו של דבר, שתנאי הנביטה של אקליפטוס המקור בדרום אפריקה טובים בהרבה מאשר בישראל. בעוד שבדרום אפריקה המין נדרש להתגבר על שני חודשי יובש, שבהם משטר המים בקרקע משופר הן בשל הטמפרטורות הממוזגות יחסית והן בשל הצמידות של עונת היובש הקצרה לעונת הגשמים, בישראל הנבטים נדרשים להתמודד עם עונת יובש ארוכה, שבסופה כמות המים בקרקע נמוכה מאד, הן בשל פרק הזמן הארוך של העדר גשמים והן בשל הטמפרטורות הגבוהות במהלך הקיץ. לכן, בקייפטאון, קל יחסית לנבטי האקליפטוס לשרוד ולהתבגר, אך בישראל הנבטים מתקשים להתגבר על המכשול האקלימי ותכופות מתייבשים. כאמור, להצלחת ההתבססות של נבטי אקליפטוס המקור נדרשת קרקע לחה וחשופה. ערכי הטמפרטורה בקרקע חשופה קיצוניים הרבה יותר מאשר בקרקע מכוסה בצומח, ולכן באקלים הישראלי, תנאי הצחיחות בקרקע חשופה עשויים להיות קטלניים עבור האקליפטוס.
בהשוואת האקלים הים תיכוני בין קליפורניה (סאן פרנסיסקו) לבין ישראל (בית-דגן) מסתבר, שהתפלגות המשקעים והטמפרטורות דומה מאוד (איור 1). עם זאת, נבטי אקליפטוס המקור עשויים ליהנות ממקור מים נוסף לאורך החופים של קליפורניה שאינו קיים בישראל. חופי קליפורניה נתונים למספר רב של ימי ערפל כבד בקיץ (ראות קטנה מ- 0.8 ק"מ לבמשך 8 שעות במהלך היממה). מתוך נתונים שנמדדו לאורך רצועת החוף של קליפורניה בין הערים ארקטה (Arcata) לקרסנט (Crescent), ישנם למעלה מ- 45 ימי ערפל במהלך החודשים יוני-אוקטובר (Dawson 1998). תופעת הערפל ידועה כגורם המשפר את יכולת ההישרדות של נבטים במהלך הקיץ במספר מינים, כמו לדוגמה בסקוויה נאה (Sequoia sempervirens) (Dawson 1998). תופעת הערפל מביאה הן לשיפור משטר המים בקרקע על-ידי תוספת ישירה של משקעים והן לשימור תכולת המים על-ידי הפחתה בעוצמת האוופוטרנספירציה. חופת העצים לוכדת מהערפל כמויות משמעותיות של מים הניגרות אל הקרקע ומחלחלות לתוכה. בחישוב תרומת הערפל לכמות המים ממנה נהנים עצי אקליפטוס גלובולוס בסן פרנסיסקו נמצא, כי ימי הערפל במהלך הקיץ תורמים כמות שוות ערך ל- 420 מ"מ, הזהה לתרומת ימי הגשם במהלך החורף (Cal-IPC 2015). תרומת מים זו לשכבת הקרקע הרדודה עשויה להיות חיונית מאוד להתבססות נבטי אקליפטוס גלובולוס. חופת העצים המצלה על הקרקע ממתנת אף היא את האוופוטרנספירציה, ובכך מסייעת לשמר את תכולת המים בשכבת הקרקע העליונה. תכולת מים גבוהה בשכבת הקרקע העליונה משפרת את תהליכי ההטמעה בצמח, וכתוצאה מכך מביאה לגידול מהיר יחסית של הנבטים בהשוואה לגידול בתנאי טמפרטורות דומים על קרקע יבשה. כלומר, תופעת הערפל לאורך חופי קליפורניה עשויה לקדם התפשטות בזחילה של אקליפטוס גלובולוס משולי החורשות, והתבססות עומדים צפופים מחוץ להן, בנוסף לנוכחות עצים בוגרים המשפרים את תנאי ההישרדות של הנבטים בסביבתם הקרובה. ואכן מדווח, כי חריגת עצי אקליפטוס גלובולוס מהחורשות הנטועות מתרחשת בעיקר באזורים הנתונים תחת השפעת ערפל (Cal-IPC, 2015).
העונה היבשה בקאדיז בספרד, קצרה אף היא בהשוואה לישראל, ומסתכמת בשני חודשים יבשים שבהם כמות המשקעים הממוצעת נמוכה מ- 5 מ"מ, ושני חודשים נוספים שבהם כמות המשקעים היא בטווח של 15-10 מ"מ. אבל בעוד שבחודש מאי בישראל כמות המשקעים הממוצעת עומדת על 5 מ"מ, כמות המשקעים בקאדיז היא 35 מ"מ. כיוון שערכי הטמפרטורה דומים, אך גבוהים במעט בישראל, המסקנה היא שגם בהשוואה לקאדיז הנבטים בישראל צריכים לשרוד עונה יבשה צחיחה יותר ולמשך זמן ארוך יותר.
בסיס הידע ששימש להגדרת אקליפטוס המקור כמין פולש – וממצאים ראשוניים של סקרי שדה
אקליפטוס המקור נכלל ברשימת הצמחים הפולשים בישראל (דופור-דרור, 2010) ללא הצגת נתונים כמותיים על קצב התפשטותו. דופור-דרור (2010, 2015) מציג גישה להגדרה כמותית של הצמחים הפולשים המבוססת על פרסומים ידועים בתחום האקולוגיה של הפלישה (Richardson et al., 2000, Pyšek et al., 2004). הגדרה זו קובעת כי קצב הפלישה נעשה ב"שיעור שעולה על 100 מטר ממקור הזרעים תוך 50 שנה, או לפחות שישה מטרים תוך שלוש שנים אצל צמחים בעלי קנה שורש זוחל" (מתוך דופור-דרור, 2016). אולם מסתבר, שסיווג המינים ברשימת הצמחים הפולשים בפועל בספרו של דופור-דרור, לא התבסס על קריטריונים כמותיים אלא על קריטריונים איכותניים. למעשה חסרים נתונים אודות קצבי ההתפשטות של כל המינים הזרים שסווגו כפולשים, ובכלל זה גם אקליפטוס המקור. למיטב ידיעתנו, טרם נעשתה עבודה על קצבי ההתפשטות של אקליפטוס המקור התומכת בהגדרתו כפולש בישראל, ומן הראוי לקדם מחקר בנושא. יתרה מזאת, מרחק ההפצה עליו מסתמכת הגדרתו של דופור-דרור, אינו מהווה לכשעצמו קריטריון עיקרי להגדרת צמחים פולשים. Pysek וחוב' (2004) מדגישים, כי צמחים פולשים יוצרים לרוב אוכלוסיות צפופות במרחק מצמח האם. ההתפשטות והצפיפות הם הגורמים ההופכים מין פולש לפוגעני (Williamson and Fitter, 1996). ואכן, התפשטות מין על פני שטח בדגם כתמי, חמורה יותר מהתפשטותו בדגם קווי (לאורך תעלה, או צידי כביש) לאורך אותו מרחק הפצה. כמו כן, התפשטות של אלפי פרטים לאורך מרחק מסויים, חמורה בהרבה מהתפשטותם של פרטים בודדים לאורך אותו המרחק.
בתצפיות שנעשו ברחבי הארץ באתרים שבהם אקליפטוס המקור ניטע לצרכי ייעור או יער משקי, נמצאו זריעים (עצים שמקורם בריבוי עצמאי מזרעים של צמחי האם) רק באתרים בודדים שרובם היו בתוך החלקות הנטועות. דופור-דרור (2010) מציין מספר מוקדים בהם ישנה פלישה של אקליפטוס המקור בשטחים טבעיים: "גן לאומי חולות קדימה, שמורת השיטה המלבינה באשדוד, גן לאומי הרי יהודה (גדות נחל כסלון), שמורת נחל שורק, שמורת לימן, שמורת כרי נעמן, שמורת נחל יבנאל, שמורת נחל תבור ושמורת תל-דן". כמו כן, דווח ש"התבססות ויצירת סבך של אקליפטוס המקור" קיימת בערוצי הנחלים משושים ודליות בגולן (דופור-דרור, מסמך חוות דעת עבור מועצה אזורית גולן, 2015).
ערכנו תצפיות ראשוניות בחלק מהאתרים האלו והממצאים מצביעים על פער גדול בין הדיווחים בספרות לבין המצב בפועל. בשמורת תל-דן, ששוכנת באזור נביעות והקרקע בה עתירת מים, לא נמצאו זריעים במרחק הגדול מ- 30 מ' מצמחי האם. באזור גן לאומי הרי יהודה, בו ניטעו עצי אקליפטוס רבים לאורך ערוצים לרבות גדות נחל כסלון, לא נמצאו זריעים במרחק מצמחי האם לאורך גדות הנחל והערוצים. שמורת לימן הינה שמורה קטנה השוכנת על גבעת כורכר בין הישוב לימן לצומת-בצת. הגבעה מוקפת בשדרות אקליפטוס מכל עבריה, ולמרות זאת, לא נמצא אף זריע אחד במרחק מצמחי האם בתוך השמורה. שמורת כרי נעמן שוכנת על תשתית לחה בצמוד לגדות הנעמן, והיא גובלת בשדרות אקליפטוס המקור ממזרח וממערב. בשדרה המזרחית נראית התרחבות מסוימת של קבוצת העצים הנטועה לכיוון השמורה (איור 2), אולם זריעים בודדים בלבד נמצאו במרחק רב מגבולות העצים הנטועים. ראוי לציין, שלאורך שטחי הצפה של גדות נחל הנעמן, נצפתה תופעה של ריבוי זריעים והתבססותם מתחת לחופות העצים, ביניהם ובסמוך להם.

איור 2. זריעים של אקליפטוס המקור גדלים בין חופות העצים של צמחי האם ובסמוך אליהם. בתמונה נראים זריעים על הגדה של בריכת הדגים, בסמוך לשדרת העצים הנטועה לאורך נחל הנעמן. צילם: עודד כהן ©
תופעה דומה נצפתה גם לאורך שדרת העצים הנטועים על גדות הקישון בקטע צומת התשבי – כביש 4 (איור 3).

איור 3. נבט גדול יחסית של אקליפטוס המקור הגדל בתנאים מיוחדים של פשט ההצפה במורד נחל קישון. במרבית אפיקי הנחלים בארץ, שבהם העונה היבשה היא לפחות חמישה חודשים, הנבטים אינם שורדים לרוב. בפשטי ההצפות, נבטי אלקיפטוס המקור נהנים הן ממשטר מים משופר יחסית במהלך הקיץ והן מהסרת הגורם התחרותי על ידי השטפונות. צילם: עודד כהן ©
בשמורת השיטה המלבינה, השוכנת על הדיונות החופיות בצפונה של אשדוד, נטועים עצי אקליפטוס בשתי חורשות (50X50 מ' בקירוב כל אחת) ולאורך שביל כורכר החוצה את השמורה. אנו מצאנו שני זריעים בלבד בכל השמורה המרוחקים מצמחי האם. עצים רבים ניטעו בשמורת נחל שורק מצפון (חורשת הצנחנים) ומדרום לכביש פלמחים. זריעים רבים אמנם נראו גדלים על גדות הנחל בצל העצים וביניהם, אך לא נצפו כלל זריעים במרחק מצמחי האם. שמורת נחל יבנאל ממוקמת בפתחת עמק כנרות, ובה עובר נחל יבנאל הזורם כל השנה. עצי אקליפטוס רבים נטועים באגן הניקוז במעלה הנחל. נמצאו זריעים בודדים בלבד הפזורים בתוך סבך הגדות במרחק מצמחי האם ובין חורשות עצים הנטועות בתוך הנחל. הדיווח על התפשטות זריעים בנחל דליות מתייחס לקטע שממזרח למחצבת כורסי וממערב לשמורה הסגורה של גמלא. האזור שוכן על תשתית מניפת סחף. בקטע זה השופע מעיינות, נחל דליות מתחתר ויוצר ערוץ זרימה איתן, שלצידיו רשת של ערוצי זרימה שיטפוניים. קבוצת עצי אקליפטוס המקור אדירי מימדים נטועה בתוך הנחל, וקבוצה נוספת על גדת הנחל הדרומית. עשרות זריעי אקליפטוס, שמקורם כנראה בעיקר בקבוצת העצים במרכז הנחל, מתפשטים במרחק של 300 מ' בקירוב במורד הנחל, בעיקר לאורך ערוצי זרימה שיטפוניים איור 4).

איור 4. התפשטות זריעים של אקליפטוס המקור בנחל דליות. בתמונה נראים עצים בוגרים שניטעו בתקופה הסורית בנחל (מימין) ובמרחק ממנו (ברקע). בעוד שקבוצת העצים המרוחקת מהנחל שומרת על גבולותיה, בנחל נראית תופעה חריגה של התפשטות עשרות זריעים עד למרחק של מאות מטרים מהחורשה הנטועה. צילם: עודד כהן ©
תצפיות שדה בערוץ נחל משושים נעשו בקטע שבין כביש בית המכס – חד-נס לבין בריכת המשושים. קבוצות רבות של עצי אקליפטוס ניטעו באגן הניקוז במעלה הנחל, חלקן עוד בתקופה הסורית. כמו כן, חורשות אקליפטוס ניטעו גם במורדות הפונים אל הנחל ולאורך קטעים ממנו. אף על פי כן, זריעים בודדים בלבד (בחלקם ספק עצים נטועים) נצפו פזורים בסבך של הצומח על גדות הנחל ובמרחק מעצים בוגרים. נחל תבור מתחתר בשכבות הבזלת של הגליל המזרחי. דיווחים של רט"ג (ד. קפלן וי. פורת, מידע בעל פה) מלמדים על התפשטות אלפי זריעים של אקליפטוס המקור במרחק רב מצמח האם, לאורך הערוץ התחתון. רט"ג טיפלו בהתפשטות זו בהצלחה, אך בעלות גבוהה (ע. דולב, מידע בעל פה). כיוון שבעקבות טיפולם השטח השתנה ללא היכר, הסתמכנו על דיווחיהם.
לאור המידע שאספנו עד כה לגבי האתרים שבהם נמצאו זריעים החורגים מגבולות החלקה בה ניטע אקליפטוס המקור, ניתן להגיע למסקנות הבאות:
- ברוב אזורי הארץ לא מוצאים תופעה של ריבוי זריעים צפוף במרחק מעצים נטועים. מצב זה שונה באופן מהותי מדרום-אפריקה, בה מוצאים באזורים נרחבים אלפי זריעים הכובשים שטחים טבעיים ודוחקים את הצומח הטבעי. כמו כן, מצב זה שונה בתכלית מפלישה של מינים המוכרים כפולשים, כמו שיטה כחלחלה (Acacia saligna) ופרקינסוניה שיכנית (Parkinsonia aculeata), המתפשטים במספר רב של מוקדים בישראל ומעמידים אוכלוסיות צפופות במוקדים החדשים.
- בכל נחלי האכזב בישראל, בהם האפיק יבש במשך חמישה חודשים לפחות, התופעת של ריבוי מזריעים של אקליפטוס המקור מזריעים היא חריגה. מכשול ההישרדות בשלבי הנביטה, מהווה כנראה גורם מגביל להתנחלות של המין בישראל. אפשר לשער, שמכשול ההישרדות קשור הן בגורמים אביוטיים, בעיקר קיץ ארוך ויבש, והן בהתנגדות ביוטית של הצומח המקומי. מעבר להשפעתה של העונה היבשה הארוכה בישראל, יש להדגיש שאקליפטוס המקור נובט בסוף החורף או באביב המוקדם, תקופה בה קיים גם צומח טבעי רב המתחרה עם אקליפטוס המקור על המים. אפילו נבטים של שיטה כחלחלה נצפו מתייבשים בשטחים בהם קיימת אוכלוסיה צפופה של דגניים בתקופה זו.
- זריעים מצויים בדרך כלל כמעט אך ורק באפיקי נחלים לחים, שקעים לחים ותעלות, שבהם הלחות בקרקע נשמרת גם בחודשי הקיץ. לחות זו היא כנראה המאפשרת לנבטים לעבור קיץ ראשון שחון לחלוטין באקלים הים-תיכוני האופייני לחלקים נרחבים בישראל, השונה באופן מהותי מהארצות שבהן האקליפטוס הפך לעץ פולש. אתרים לחים באפיקי נחלים, תעלות ושקעים במהלך הקיץ מצויים בעיקר בצפון ישראל ונדירים יותר בשדרת ההר המרכזית. בנחל רפאים ובנחל שורק לדוגמה, מצויים עשרות זריעי אקליפטוס בשל הביוב שהוזרם בהם במשך שנים רבות (א. שמידע, מידע בעל-פה). הזריעים הצעירים זקוקים ללחות בתקופת הקיץ, אך לאחר שהתבססו והעמיקו שורש הם יכולים להמשיך לגדול באפיקים יבשים. דוגמה לכך הוא חלקו העליון של נחל רפאים, בו באזור גן החיות התנכי גדלים 7 זריעים בגודל של מעל ל- 6 מטר. בשנים האחרונות האפיק יבש, אך בעבר הוזרם בו הביוב של ירושלים המערבית (א. שמידע, דיווח בע"פ).
דיון מחודש במצב העכשווי וברמת האיום של אקליפטוס המקור בישראל
אקליפטוס המקור ניטע בהיקף נרחב בבתי גידול רבים בישראל, כבר למעלה מ- 130 שנה. מהדיווחים על התפשטות ספונטנית של עומדים צפופים במרחק מצמחי האם, עולה שהתפשטות זו היא קטנה ביותר ביחס להיסטורית הנטיעה והיקפה, ואינה מצדיקה את הגדרת המין כפולש, לא לפי הקריטריונים המקובלים בתחום (Pyšek et al., 2004) ולא לפי הקריטריונים העדכניים שהוצעו לגבי ישראל על ידי כהן וריוב (2015). להלן מוצגת התייחסותנו לסיבות בשלן הוגדרה רמת האיום של אקליפטוס המקור בישראל כגבוהה (דופור-דרור, 2010):
- מאפייני הנישה האקולוגית של אקליפטוס המקור בתחום התפוצה הטבעי. גודל תחום התפוצה באוסטרליה אינו מלמד על הסתגלנות לתנאי האקלים השונים, שכן באקלים יובשני ויובשני למחצה בארץ זו, הצמח תופס נישה צרה מאוד, לאורך צידי נחלים ושטחי הצפה בלבד, והתפוצה הרחק ממקורות מים מעטה מאוד. באותה המידה, לא יהיה נכון להסיק משטח התפוצה של הנרקיס המצוי בישראל, שהמין מותאם לאקלים היובשני של דימונה, שכן הנרקיס, כמו צמחים ים תיכוניים אחרים הגדלים במדבר, מוצא מפלט ומתחמק מן האקלים היובשני בכיסי קרקע מוצפים למרגלות משטחי סלע.
- היקף הפלישה העולמי. אקליפטוס המקור ניטע בעשרות ארצות בעולם. ישנם דיווחים, שהמין התאזרח במספר רב של ארצות, בהן הפיליפינים, בנגלדש, פקיסטן, הוואי, פלורידה, קליפורינה, ספרד ודרום אפריקה. אולם מכל הארצות המדווחות, המין התקדם למעמד של צמח פולש רק בדרום אפריקה, שם הוא מתפשט במספר רב של מוקדים צפופים ובמידה מסוימת גם בחצי האי האיברי ובקליפורניה. בהקשר לכך נמצא לאחרונה, שנתוני שטח התפוצה הפוטנציאלי העולמי של מינים מהווים אינדיקטור טוב יותר למידת פלשנותם מאשר שטח התפוצה הטבעי (Higgins and Richardson, 2014). המחקר התבסס על מאגר נתונים לגבי 749 מינים של אקליפטוס ושיטה שמקורם באוסטרליה, שניטעו במקומות שונים בעולם.
- היקף הפלישה בישראל. זריעים גדלים לרוב בתוך חורשות נטועות ובסמוך להן. למיטב ידיעתנו קיים רק אתר אחד שבו התפתח עומד צפוף וגדול במרחק מצמחי אם והוא נחל תבור. תופעת התבססות והצטופפות זריעים סביב עצים בוגרים ובסמוך אליהם נראית בעיקר לאורך פשטי הצפה, אך קצב ההתפשטות של זריעים וקצב הצטופפותם (לשדרות ולקבוצות עצים) הוא איטי וניתן לבקרה.
- התאקלמות יערות אקליפטוס במגוון בתי הגידול בארץ. העובדה, שאקליפטוס המקור גדל במגוון בתי גידול באקלים הים תיכוני ובאקלים הצחיח למחצה בישראל, אין בה כדי לרמז על פלשנות המין. העצים הנטועים ביערות צלחו את מכשול ההישרדות בשלב הנביטה במשתלה ולא בשטח. כמו כן, השתילים טופלו באינטנסיביות בשנים הראשונות לאחר הנטיעה, כולל מתן השקיה. מלבד חשיבות הגורם האקלימי, אקליפטוס המקור רגיש יחסית לגיר, שהינו נפוץ מאוד בקרקעות ישראל. העץ סובל מכלורוזה קשה ולעתים אף תמותה בקרקעות בעלות תכולת גיר כללי גבוהה, בעיקר בקרקעות חרסיתיות. רגישות זו מגבילה את גידול אקליפטוס המקור לקרקעות בעלות רמת גיר נמוכה יחסית. כמו כן, אקליפטוס המקור מתאים לאזור הים תיכוני בלבד. ניתן לראות שעצים שניטעו באזור סמי ארידי (צפון הנגב) מתנוונים בהדרגה (יתכן שתורמת לכך גם הקרקע בעלת תכולת הגיר הגבוהה). לכן בצפון הנגב ניטע תת מין של אקליפטוס המקור, שהוא עמיד יותר ליובש (E. camaldulensis subsp. subcinerea =arida).
-
השפעות סביבתיות. ראשית יש לציין שההשפעות הסביבתיות אינן מהוות קריטריון מתאים להגדרת צמחים זרים כפולשים בעולם (Valery et al., 2008) ובישראל (כהן וריוב, 2015). זאת כיוון, שתהליך הפלישה הביולוגית הוא בראש ובראשונה תהליך ביו-דמוגרפי הקשור בהתפשטות של המין הזר בסביבה החדשה. לכן, הקריטריונים להגדרת מין פולש הינם קריטריונים ביו-דגמוגרפיים הקשורים במאפייני ההתפשטות של המין הזר ונמדדים על-ידי קצב התפשטות המין, היקף השטחים המאוכלסים בו וצפיפות הפרטים בהם (כהן וריוב, 2015). ההשפעה הסביבתית של המין, נגזרת מהתפשטותו. כמו כן, קשה לכמתה באופן שאינו תלוי בעיני המתבונן. לכן, עצם ההשפעה על הסביבה אינה מהווה קריטריון להגדרת מינים פולשים, אלא תורמת להערכת הסיכון הטמון בהם וזאת אך ורק תוך התחשבות במידת התפשטותם. דעה זו מעוגנת גם בהגדרה של מינים פולשים על ידי הארגון העולמי לשמירת טבע למינים פולשים (IUCN, 2016). הארגון מגדיר את המינים הפולשים ככאלו שהגיעו לסביבה שלא יועדה להם מראש ומחוץ לתחום תפוצתם הטבעי, העמידו בה אוכלוסיה בת-קיימא, והתפשטו ממנה לבתי גידול נוספים. להתפשטות זו, ישנן השלכות שליליות על המערכות האקולוגיות המקומיות והמינים המתקיימים בהן. ההשפעות הסביבתיות של אקליפטוס המקור נחקרו במספר מאמרים. במחקרים שפורסמו לאחרונה לגבי השפעת המין על מדדי עושר ומגוון הצומח והחי בדרום אפריקה (Terrerai et al., 2013) ובישראל (Herrmann et al., 2015), נמצא הבדל בהרכב חברות החי והצומח בין יערות נטועים לשטחים הסמוכים אליהם, המתבטאים בהפחתה של המינים המקומיים וכניסה של מינים מזדמנים. אולם בניגוד לצמחים פולשים אחרים, שהשפעתם על הסביבה היא בלתי הפיכה, או הפיכה אבל בהשקעת משאבים רבה מאוד, ההשפעות של האקליפטוס על התכונות הכימיות (Tererai and Gaertner, 2014) וההידרולוגיות של הקרקע הן מעטות ואינן ארוכות טווח (Ruwanza, 2013). לכך יש להוסיף, שאקליפטוס המקור אינו יוצר בנק זרעים בקרקע בדומה לשיטה הכחלחלה, וכריתה והדברה של הגדמים עשויות להביא לשיקום הצומח הטבעי בקלות יחסית. ואכן, כריתה והדברה של עצי אקליפטוס המקור באמצעות התכשיר "שוטגן" (Shotgun) הביאו לתוצאות טובות בשמורת עינות גבתון, ובהעדר התחדשות של האקליפטוס מבנק זרעים, ניכר כי הצומח הטבעי התאושש בשטחים המטופלים (נתונים שלנו).
סיכום והמלצות
לפי המידע שיש בידינו כיום, המבוסס על סקירת ספרות, דיווחים וממצאים ראשוניים של סקרי שדה, אקליפטוס המקור הוא מין שהתאזרח בישראל (כהן וריוב, 2015), כלומר מין שמתרבה עצמאית ביערות נטועים ובסמוך להם, בעיקר לאורך צידי תעלות ונחלים, ובתנאים מסוימים גם במרחק מצמח האם. כמות הזריעים ביערות הנטועים קטנה מאד, והופעת זריעים במרחק מצמח האם נדירה יחסית להיקף הנטיעות, ואינה מצדיקה את הגדרתו כפולש. לדעתנו, כל עוד לא הוכח אחרת, אין בדיווחים הקיימים כדי להצדיק את הכללת אקליפטוס המקור ברשימת המינים הפולשים בישראל, שכן המין אינו מתרבה במספר רב של מוקדים ולא מעמיד בהם אוכלוסיות צפופות. הסבירות לפלישה עתידית של אקליפטוס המקור נראית לנו נמוכה מאוד בשל הסיבות הבאות: האקליפטוס כסוג מציג תכונות פלישה נחותות בהשוואה לסוגי עצים ושיחים פולשים; שטח הפלישה העולמי של אקליפטוס המקור קטן מאוד ביחס להיקף הנטיעות הגדול מאוד שלו. הקיץ הארוך והיבש בישראל נראה כגורם מגביל המונע מהנבטים להתגבר על מכשול ההישרדות במוקדים חדשים ובמרחק מצמח האם. באשר להשפעות הסביבתיות, נטיעות אקליפטוס אכן משנות את המבנה והתפקוד של המערכת האקולוגית, אך ההשפעות כאמור אינן קריטריון העומד בפני עצמו להגדרת מינים זרים כפולשים. יתרה מזאת, ההשפעות של אקליפטוס המקור על הסביבה הן הפיכות. דיווחים מהספרות וכמו גם ניסיוננו בפעולות שיקום בישראל מלמדים, שניתן לשקם את הצומח הטבעי לאחר כריתה והדברת העצים ללא צורך בהתערבות יזומה לשינוי תכונות הקרקע ובנק הזרעים.
אנו מציעים להרחיב את המחקר להבנה של דינמיקת האוכלוסיות של אקליפטוס המקור במגוון בתי הגידול של ישראל שיכלול התייחסות ליחסי הגומלין בין הצמח לסביבה. מוסכם על הכל, שקצבי ההתפשטות של אקליפטוס המקור קטנים באופן משמעותי מאלו של צמחים פולשים המסכנים את המערכות האקולוגיות. לאור העובדה שקצב ההתפשטות של מין זה איטי והשפעותיו הן הפיכות, נראה שיש מקום לערוך מחקר מבוסס בתחום לפני קבלת החלטות ממשקיות, ולא להיפך.
לאור לחצי הפיתוח הגבוהים בישראל ומגוון האיומים לשמירת הטבע בנוסף לפלישה הביולוגית, אין מקום לריבוי ספונטני של צמחים זרים, ולכן מומלץ לכרות ולהדביר את כל זריעי אקליפטוס המקור בתוך שטחי הנטיעות ומחוצה להן בשטחים הפתוחים בכלל ובשמורות הטבע בפרט, ולהימנע מנטיעות נוספות לאורך נחלים. טיפול בהתחדשויות מזריעים נראה לנו כפתרון יעיל מכל הבחינות בהשוואה לכריתה של עצים בוגרים. פעולות לוויסות של האוכלוסיה הן מפתח לפתרון המחלוקת בין שימור המגוון הביולוגי מחד, לבין שימור ערכי המורשת ושירותי המערכת האקולוגית של אקליפטוס המקור מאידך.
ספרות:
גליקסמן א 2007 בחינת השימוש ברסק אקליפטוס ככלי ידידותי לסביבה בהדברת עשבים. עבודת גמר, האוניברסיטה העברית בירושלים.
דופור-דרור ז מ 2010 הצמחים הפולשים בישראל. רשות הטבע והגנים הלאומיים, המשרד להגנת הסביבה. הוצאת אחווה, ירושלים.
דופור-דרור ז מ 2015 הגדרת צמחים פולשים והתייחסות למעמדו של אקליפטוס המקור כפולש אפשרי בישראל. כלנית כתב עת לצמחי ישראל . https://www.kalanit.org.il
ויינשטיין א זהר י זהר וברנד ד 2011 האקליפטוס מצמרתו ומעלה. אקליפטופ בע"מ, פתח תקווה.
רוזן ב 1991 ראשית אקלום האקליפטוסים בארץ-ישראל. קתדרה 59. הורד ב- 17.12. 2015:http://www.snunit.k12.il/heb_journals/katedra/59095.html
כהן ע וריוב י 2015 קריטריונים להגדרת צמחים פולשים בישראל ולהערכת הסיכון. כלנית 2. https://www.kalanit.org.il/?p=4680
רגיל ב 2002 נוכחות חומרים מעכבי נביטה בעלי אקליפטוס והשפעתם על זרעים וצמחים חיים. עבודת גמר, בית ספר אזורי מקיף ע"ש י.ח. ברנר, הפקולטה לחקלאות, רחובות.
———————————————————————————————————————
Alpert P Bone E and Holzapfel C 2000 Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. Perspectives in Plant Ecology, Evolution and Systematics 3: 52–66.
Baskin CC and Baskin JM 2014 Seeds: Ecology, Biogeography, and Evolution of Dormancy. Second edition. Academic Press, San Diego, CA.
Becerra PI and Bustamante RO 2008 The effect of herbivory on seedling survival of the invasive exotic species Pinus radiata and Eucalyptus globulus in a Mediterranean ecosystem of Central Chile. Forest Ecology and Management 256:1573–1578.
Booth T 2014 Modern tree colonisers from Australia into the rest of the world. pp 304–323 in H H Prins and P L Gordon (eds.). Invasion Biology and Ecological Theory: Insights from a Continent in Transformation. Cambridge University Press. Cambridge, United Kingdom.
Booth TH 2012 Eucalypts and their potential for invasiveness particularly in frost-prone regions. International Journal of Forestry Research 2012 :1–7.
Cal-IPC 2015 Assessment of Tasmanian blue gum (Eucalyptus globulus). http://www.cal-ipc.org/ip/inventory/pdf/Eucalyptus_globulus_PAF_2015March.pdf.
Callaham MA Stanturf JA Hammond WJ Rockwood DL et al. 2013 Survey to evaluate escape of Eucalyptus spp. seedlings from plantations in Southeastern USA. International Journal of Forestry Research. http://cx.doi.org/10.1155/2013/946374.
Callaway RM and Aschehoug ET 2000 Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 290: 521–523.
Calviño-Cancela M and Rubido-Bará M 2013 Invasive potential of Eucalyptus globulus: Seed dispersal, seedling recruitment and survival in habitats surrounding plantations. Forest Ecology and Management 305: 129–137.
Chambers JC and Macmahon JA 1994 A day in the life of a seed – movements and fates of seeds and their implications for natural and managed systems. Annual Review of Ecology and Systematics 25: 263–292.
Cremer KW 1977 Distance of seed dispersal in eucalypts estimated from seed weights. Australian Forest Research 7: 225-228.
Dana ED Sanz-Elorza M Vivoas M and Sobrino S 2005 Especies vegetales invasoras en Andalusa. Coneejeria de Medio Ambiente, Junta de Andalucia, Sevilla.
Dawson TE 1998 Fog in the California redwood forest: ecosystem inputs and use by plants. Oecologia 117: 476–485.
Del Moral R and CH Muller 1970 The allelopathic effect of Eucalyptus camaldulensis. The American Midland Naturalist 83: 254-282.
Díez J 2005 Invasion biology of Australian ectomycorrhizal fungi introduced with eucalypt plantations into the Iberian Peninsula. Biological Invasions 7: 3–15.
Di Stefano J 2001 River red gum (Eucalyptus camaldulensis): a review of ecosystem processes, seedling regeneration and silvicultural practice. Australian Forestry 65: 14–22.
Domingues De Almeida J and Freitas H 2006 Exotic naturalized flora of continental Portugal – A reassessment. Botanica Complutensis 30: 117–130.
Dufour-Dror JM 2012 Alien Invasive Plants in Israel. The Middle East Nature Conservation Promotion Association, Ahva publishing: 213.
Forsyth GD Richardson P Brown and Van Wilgen B 2004 A rapid assesment of the invasive status of Eucalyptus species in two South African provinces. South African Journal of Science 100: 75–77.
Gassó NW Thuiller W and Pino J 2012 Potential distribution range of invasive plant species in Spain. Nobiota 12: 25–40.
Gordon DR Tancig KJ Onderdonk DA and Gantz CA 2011 Assessing the invasive potential of biofuel species proposed for Florida and the United States using the Australian Weed Risk Assessment. Biomass and Bioenergy 35: 74–79.
Herrmann JI Opatovsky Y Lubin T Pluess E et al. 2015 Effects of non-native Eucalyptus plantations on epigeal spider communities in the northern Negev desert, Israel. The Journal of Arcachnology 43: 101–106.
Higgins SI and DM Richardson 2014 Invasive plants have broader physiological niches. Proceedings of the National Academy of Sciences of the United States of America 111: 10610–10614.
Lake J Richardson DM and Pysek P 2004 Invasion success of exotic plants in natural ecosystems: the role of disturbance, plant attributes and freedom from herbivores. Biological Conservation 117: 215–226.
Lorentz K A and Minogue PJ 2015 Exotic Eucalyptus plantations in the southeastern US: risk assessment, management and policy approaches. Biological Invasions 17: 1581–1593.
Lorentz K A and Minogue PJ 2015 Potential Invasiveness for Eucalyptus Species in Florida. Invasive Plant Science and Management 8: 90–97.
May F and Ash J 1990 An assessment of the allelopathic potential of Eucalyptus. Australian Journal of Botany 38:245-254.
Moles A and Westoby M 2004 Seedling survival and seed size: a synthesis of the literature. Journal of Ecology 92: 372-383.
O'Dowd DJ and Gill AM 1984 Predator satiation and site alteration: mass reproduction of alpine ash (Eucalyptus delegatensis) following fire in southeastern Australia. Ecology 65: 1052-1066.
Pallewatta N Reaser JK and Gutierrez AT 2003 Invasive species in South-Southeast Asia: national reports and directory of resources.
Pedrol N Gonzalez L and Reigosa MJ 2006 Allelopathy and abiotic stress. in: Reigosa M J Pedrol N and Gonzalez L (eds). Allelopathy: A Physilogical Process with Ecological Implications. Springer, Dordrecht, The Netherlands. pp. 171-209.
Potts BM and Wiltshire J 1997 Eucalypt genetics and geneogology. in Williams J and Woinarski J (eds.) Eucalypt Ecology: Individuals to Ecosystems. Cambridge University Press, Cambrige, United Kingdom. pp. 56–91.
Protasov A La Salle J Blumberg D Brand D et al. 2007 Biology, revised taxonomy and impact on host plants of Ophelimus maskelli, an invasive gall inducer on Eucalyptus spp. in the Mediterranean area. Phytoparasitica 35: 50–76.
Pysek P Richardson DM Rejmanek M Webster GL et al. 2004 Alien plants in checklists and floras: towards better communication between taxonomists and ecologists.Taxon 53(1): 131-143.
Rejmánek M and Richardson DM 2011 Eucalypts. in Simberloff D and Rejmanek M (eds.) Encyclopedia of Biological Invasions. University of California Press, Berkeley and Los Angeles, CA. pp. 203–209.
Richardson D Pysek MP Rejmánek M Barbour MG et al. 2000 Naturalization and invasion of alien plants: concepts and definitions. Diversity and Distributions 6: 93–107.
Ritter M and Yost J 2009 Diversity, Reproduction, and Potential for Invasiveness of Eucalyptus in California. Madroño 56: 155–167.
Roberts J and Marston F 2011 Water regime of wetland and floodplain plants: a source book for the Murray-Daling Basin, National Water Commision, Canberra, Australia.
Ruwanza S Gaertner M Richardson DM and Esler KJ 2013 Soil water repellency in riparian systems invaded by Eucalyptus camaldulensis: A restoration perspective from the Western Cape Province, South Africa. Geoderma 200-201: 9–17.
Ruwanza S Gaertner M Esler KJ and Richardson DM 2015 Allelopathic effects of invasive Eucalyptus camaldulensis on germination and early growth of four native species in the Western Cape, South Africa. Southern Forests 77: 91–105.
Tererai F Gaertner M Jacobs SM and Richardson DM 2013 Eucalyptus invasions in riparian forests: Effects on native vegetation community diversity, stand structure and composition. Forest Ecology and Management 297:84–93.
Tererai F Gaertner M Jacobs SM and Richardson DM 2015 Eucalyptus camaldulensis invasion in riparaian zones reveals few significant effects on soil physico-chemical properties. River Research and Applications 31: 590-605.
Thompson K 1987 Seeds and seed banks. New Phytologist 106: 23-34.
Traba JF Ascarate and Peco B 2004 From what depth do seeds emerge? A soil seed bank experiment with Medierranean grassland species. Seed Science Research 4: 297-303.
Turnbull JW 1999 Eucalypt plantations. New Forests 17:37–52.
Valéry LH Fritz JC Lefeuvre D and Simberloff D 2008 In search of a real definition of the biological invasion phenomenon itself. Biological Invasions 10: 1345–1351.
von der Lippe M and Kowarik I 2007 Long-distance dispersal of plants by vehicles as a driver of plant invasions. Conservation Biology 21: 986–996.
Williamson M and Fitter A 1996 The varying success of invaders. Ecology 77: 1661–1666.
Wilson EO 2003 The Future of Life. Abacus, London, Uniteed Kingdom, pp. 230.
Zedler JB and Kercher S 2004 Causes and consequences of invasive plants in wetlands: opportunities, opportunists, and outcomes. Critical Reviews in Plant Sciences 23: 431–452.
===========================================
כל הזכויות שמורות ל"כלנית" ©
ציטוט: כהן ע וריוב י 2016 האם אקליפטוס המקור הוא מין פולש בישראל? כתב-עת "כלנית" מספר 3.
https://www.kalanit.org.il/?p=6628&preview=true
עוד בכלנית על אקליפטוס המקור
קריטריונים להגדרת צמחים פולשים בישראל ולהערכת הסיכון
הגדרת צמחים פולשים והתייחסות למעמדו של אקליפטוס המקור כפולש אפשרי בישראל










