אמוץ דפני, החוג לביולוגיה אבולוציונית, אוניברסיטת חיפה adafni@research.haifa.ac.il
חגי צהרי, החוג לביולוגיה אבולוציונית, אוניברסיטת חיפה hagai11@gmail.com|
רחל בן-שלמה החוג לביולוגיה וסביבה, אוניברסיטת חיפה, אורנים ekly@research.haifa.ac.il
ניקולס ווריקן, Agroecology & Pollination Group, Landscape Ecology & Plant Production Systems, Université Libre de Bruxelles, Belgium Nicolas.vereecken@ulb.ac.be
גידי נאמן, החוג לביולוגיה וסביבה, אוניברסיטת חיפה, אורנים gneeman@gmail.com
מאמר זה מבוסס על המאמר המדעי:
Dafni A Tzohari H Ben-Shlomo R Vereecken N et al. 2020 Flower colour polymorphism, pollination modes, breeding system and gene flow in Anemone coronaria. Plants 9 (3) :397 https://www.mdpi.com/2223-7747/9/3/397.
תקציר: כלנית מצויה Anemone coronaria ממשפחת הנוריתיים Ranunculaceae מאופיינת בפרחים אדומים, לבנים, כחולים, בגוונים שונים של סגול-ארגמן או וורוד ותכונה זו היא תורשתית. פולימורפיזם (רבגוניות) קיצוני כזה של צבעי פרחים במסגרת מין אחד היא תופעה נדירה יחסית. רבגוניות כזו עשויה להיות תוצאה של ברירה טבעית על-ידי המאביקים, על-ידי גורמים ביוטיים אחרים שאינם מאביקים, או על-ידי תנאי סביבה. במחקר זה בדקנו את ההרכב הגנטי ומידת הדמיון הגנטי בין עשר אוכלוסיות של כלניות בעלות פרחים אדומים לבנים וכחולים הגדלות בעשרה אתרים בישראל ובין טיפוסי הצבע השונים של כלניות ההגדלות באוכלוסיות מעורבות. כמו כן בדקנו את מנגנוני הזיווג, דרכי ההאבקה והתנהגות המאביקים בטיפוסי הצבע השונים. מצאנו שלפרחים בצבעים שונים יש החזר אור שונה שדבורי דבש מסוגלות להבדיל ביניהם. חנטת הפירות בפרחים שהיו תחת רשת שאפשרה האבקת רוח אך מנעה גישת מאביקים, הייתה דומה לזו של פרחים שהיו חשופים להאבקה טבעית על-ידי מאביקים ורוח. תוצאה זו מעידה על התפקיד המשמעותי של האבקת רוח בפרחי הכלנית. פרחי הכלנית בוקרו על-ידי חרקים שונים בעיקר דבורים וזבובים, אבל בניגוד למחקרים קודמים לא נצפו חיפושיות בפרחים. דבורי דבש, ודבורים יחידאיות Andrena sp. העדיפו באופן מובהק פרחים לבנים וכחולים על פני אדומים, זבובי רחף (Syrphidae) נמנעו גם הם מלבקר בפרחים האדומים, אך לא באופן מובהק. לכן, התנהגות המאביקים יכולה להסביר רק באופן חלקי את מגוון הצבעים בכלנית. הבדיקות הגנטיות של הרכב הDNA, לא הראו הבדלים מובהקים בין האוכלוסיות ולא בין טיפוסי הצבע האדומים, לבנים והכחולים. התוצאות מראות שבטבע קיים חילוף חופשי של גנים בין האוכלוסיות ובין טיפוסי הצבע השונים, ושהאבקת רוח אקראית יכולה להסביר את העדר ההבדלים האלה.
מבוא
פולימורפיזם (רבגוניות) בצבע פרחים מתייחס להבדלים בולטים בצבע הפרחים בין פרטים שונים באותה אוכלוסייה או בין אוכלוסיות שונות של אותו מין (Joseph & Siril 2013). פולימורפיזם יכול להיות הדרגתי ולהכיל גוונים שונים של אותו צבע (כמו ברקפת מצויה) או צבעים שונים לחלוטין (כמו בכלנית, פשתנית ססגונית ועוד). טיפוסי הצבע השונים יכולים להיות סימפטריים, כלומר לחלוק תחום תפוצה משותף, או אלופטריים, כלומר בעלי תחומי תפוצה נפרדים (and Bierzychudek 2007; Wang et al. 2016; Narbona 2017).
דבורים לומדות מהר לזהות מקור מזון על פי מיקומו, צבעו וריחו, הן נשארות נאמנות לו וחוזרות וממשיכות לבקר בו כל זמן שהוא מספק להם גמול ראוי. נאמנות כזו לטיפוס צבע מסוים והימנעות מטיפוס צבע אחר, במסגרת אותו מין של צמח עשויה לגרום לאבולוציה מתפצלת וליצירה של שני תתי מינים, או אפילו מינים נפרדים (Chittka et al. 1999). לכן הימצאותם של טיפוסי צבע שונים במין אחד דורשים תשומת לב מיוחדת. מצב מיוחד כזה של רבגוניות יכול להיגרם על-ידי העדפה סלקטיבית של מאביקים (Gegear and Laverty 2001; Jones and Reithel 2001). במקרה כזה זרימת הגנים צפויה להיות גדולה יותר בין טיפוסי צבע זהים מאשר בין טיפוסי צבע שונים ( Malerba and Nattero 2012). לפיכך השונות הגנטית בין פרטים מאותו טיפוס צבע צפויה להיות נמוכה יותר מאשר השונות בין טיפוסי צבע שונים (Brown and Clegg 1984).
הרביוורים (אוכלי צמחים) מבחינים גם הם בצבע הפרחים ועשויים להעדיף טיפוס צבע אחד על פני אחר (Frey 2004; Johnson 2008). כך הם עשויים לגרום לסלקציה לטובת טיפוס צבע מסוים שעשויה להיות שונה, או אף נוגדת את הסלקציה של המאביקים (Fineblum and Rausher 1997). כך למשל חרקים המטילים בפרחים והניזונים מהזרעים המתפתחים בשחלותיהם יעדיפו גם הם את הפרחים המועדפים על-ידי המאביקים, מכיוון שפרחים אלה צפויים ליהנות מהאבקה טובה יותר, ליצר יותר זרעים כלומר כמות מזון גדולה יותר להרביוורים (Strauss and Whittall 2006).
סמיכות פיזית בין האתרים בגנום הקובעים את צבע הפרחים לבין אלה הקובעים את העמידות לתנאי הסביבה גורמת לתאחיזה בין תכונות אלה (כלומר שבמקרים רבים הן עוברות יחד בתורשה) ולכן סלקציה לטובת אחת מהן עשויה להשפיע על האחרת (Strauss et al. 2004; Arista et al. 2013). התאמה לתנאי סביבה לאורך מפל בתנאים סביבתי יכולה גם היא לשמר טיפוסי צבע שונים באותו מין. לדוגמא, שימור צבע הפרח בשל הקשר בין אנתוצאינין (הגורם לצבע אדום וסגול בפרחים) לעמידות הצמחים ליובש, חום, קרינת UV, דלות קרקע ואפילו עמידות למזיקים (Steyn et al. 2002).
כלנית מצויה .Anemone coronaria L היא מין מזרח ים-תיכוני הגדל במגוון בתי גידול (Horovitz et al. 1975). לכל צמח 10-1 פרחים בהתאם לגילו, גודל הפקעת והתנאים המקומיים. בכלנית קיימת שונות גדולה בצבע הפרחים. יש טיפוסים עם פרחים אדומים, לבנים, כחולים ועוד כ 30 גווני צבע סגולים-ארגמניים שונים (איור 1), וצבע הפרח נקבע על-ידי הריכוז של פיגמנטים שונים (בעיקר אנתוציאנין ופלבונואידים) בעלי הכותרת (Harvey1971).
צבע הפרח בכלניות נקבע על-די תכונה תורשתית שנחקרה רבות על-ידי אורה הורוביץ ז"ל מהאוניברסיטה העברית בירושלים. הצבע נקבע לפחות על-ידי ארבעה גנים אפיסטטיים הפועלים בטור, זה אחר זה על פי הסדר הבא: 1. אלבינו (Alb) 2. סגול (Sc) 3. לבן (W) 4. כחול (B) (Horovitz 1990, Horovitz 1995; Horovitz and Zohary 1996 ). אוכלוסיות של כלניות אדומות מואבקות בעיקר על-ידי חיפושיות-פרחים, גלפיריות Glaphyridae, אך גם דבורי דבש נמצאו מבקרות ואוספות אבקה בפרחים אדומים (Dafni et al. 1990; Keasar et al. 2008). החיפושיות מוצאות בפרחים אבקה כמזון, מקלט לילה וגם אתר להזדווגות. נמצא שחומרים רעילים anemonine בעלי הפרחים והצבע האדום מספקים להם גם הגנה חלקית מטריפה על-ידי ציפורים (Keasar et al. 2010; Keasar et al 2013).
בישראל, האוכלוסיות הדרומיות של כלניות הגדלות באזור הספר, בדרום החבל הים-תיכוני, מכילות רק פרחים אדומים. אוכלוסיות אחדות בצפון (באזור עמיעד-ראש פינה) הן לבנות ובגולן גדלות אוכלוסיות שהן בעיקר בגוונים שונים של סגול-ארגמן. אולם מרבית האוכלוסיות בגליל ובכרמל הן רבגוניות ומכילות פרטים בעלי פרחים בצבעים שונים, אך גם אוכלוסיות קטנות אדומות בלבד. כלניות בעלות פרחים אדומים נמצאו עמידות לרעייה חזקה של בקר ברמת הנדיב ומספר הפרטים ירד בחלקות שבהן הופסקה רעיית הבקר (Perevolotsky et al. 2011). בניגוד לבקר, הפרחים של הכלניות האדומות נאכלים על-ידי איילים (Wallach et al. 2009).
מטרות המחקר היו: (1) בחינת האקולוגיה של האבקה בכלנית מטיפוסי צבע שונים. (2) בדיקה של השונות הגנטית בין האוכלוסיות ובין טיפוסי הצבעים השונים בתוך האוכלוסיות. (3) הצעת הסברים אפשריים לרבגוניות הקיצונית בצבע פרחי הכלניות בישראל. לשם כך ניסינו לענות על שאלות המחקר הבאות: (1) האם הדבורים המבקרות בפרחי הכלנית מסוגלות להבחין בין הצבעים השונים של פרחי הכלניות כפי שרואה אותם האדם? (2) האם מאביקים שונים מעדיפים לבקר בפרחי כלנית בצבעים שונים? (3) מה התפקיד שממלאות האבקה עצמית והאבקת רוח במנגנון הרבייה של הכלניות? (4) האם ההרכב הגנטי והשונות הגנטית מראים אפשרות של בידוד רבייתי בין האוכלוסיות ובין טיפוסי הצבע השונים בתוכן?

איור 1. פריחה של כלניות אדומות, לבנות, כחולות ובעלות צבעי ביניים, ומראה כללי של אוכלוסייה מעורבת. צילום חגי צהרי ©

חומרים ושיטות
אתרי מחקר
תצפיות בחרקים המבקרים בפרחי כלניות נערכו בשנים 2011, 2012 ו 2014 בשישה אתרים בישראל שבמרביתם נמצאו טיפוסי צבע שונים (טבלה 1, איור 2). אנליזה של צבע הפרחים נבדקה באמצעות החזר גלי האור באורכי גל שונים מפרחים אדומים לבנים וכחולים באתר מגידו. חומר צמחי לבדיקות DNA נלקח מצמחים בעלי טיפוסי צבע שונים בעשרה אתרים שונים מרחבי תחום התפוצה של הכלנית המצויה בישראל (טבלה 1). בכל אתר נדגמו צמחים משטח של כ- 1000 מ"ר.
טבלה 1. אתרי המחקר, המיקום הגאוגרפי שלהם (GPS) וצבע הפרחים באתר (אדום R, כחול B, לבן W). האתרים בהם נערכו תצפיות במבקרים בפרחים ב 2011-2012 וב 2014, האתרים מהם נלקח חומר לבדיקת DNA ופרחים לאנליזה של הצבע מסומנים ב- ✔.
| אתר המחקר | GPS N/E | צבע הפרחים | תצפיות 2011-12 | תצפיות 2014 | אנליזת DNA |
אנליזת צבע |
| 1. אלוני אבא | 32.73/35.17 | RBW | ✔ | |||
| 2. אלונים | 32.72/35.15 | RBW | ✔ | ✔ | ||
| 3. בלפוריה | 32.63/35.29 | RBW | ✔ | |||
| 4. בית לחם הגלילית* | 32.74/35.19 | RBW | ✔ | |||
| 5. בית שערים | 32.70/35.13 | RW | ✔ | |||
| 6. אוניברסיטת חיפה | 32.75/35.03 | R | ✔ | |||
| 7. להבים | 32.37/34.85 | R | ✔ | |||
| 8. מגידו | 32.59/35.23 | RBW | ✔ | ✔ | ✔ | ✔ |
| 9. הר מירון | 33.01/35.40 | RBW | ✔ | |||
| 10. נחל סנין | 32.64/35.10 | RBW | ✔ | |||
| 11. דרך נוף כרמל | 32.75/35.05 | RBW | ✔ | |||
| 12. פורה | 31.50/34.78 | R | ✔ | |||
| 13. ציפורי | 32.76/35.24 | RBW | ✔ | ✔ |
* שתי אוכלוסיות

איור 2. מיקום אתרי המחקר וציון הרכב הצבעים של הכלניות הפורחות בהם.
החזר גלי האור מהפרחים וראייתם על-ידי דבורים
למדידת ההחזר היחסי (%) של גלי האור דגמנו 15 פרחים, מכל צבע, אדומים, לבנים וכחולים, כל אחד מצמח נפרד, באתר מגידו. למדידת הקרינה החוזרת באורכי גל שונים (700-300 ננומטר בצעדים של 5 ננומטר) נבדק החלק המרכזי של עלי הכותרות של פרחי כלנית צעירים מהצבעים השונים. לבדיקה השתמשנו בספקטרופוטומטר ידני (AVASPEC-2048-USB2-UA; Avantes, Eerbeek, The Netherlands) שכויל בעזרת צבע לבן סטנדרטי (AVALIGHT-XE; Avantes). לאחר מכן, על מנת לבדוק האם דבורים מבחינות בהבדלים בין הצבעים השתמשנו בעקומת הרגישות הספקטרלית של דבורת הדבש (Apis mellifera) הידועה כמייצגת היטב גם מיני דבורים נוספים (Peitsch et al. 1992; Chittka and Kevan, 2005; Dyer et al. 2008). בהמשך שרטטנו את ההחזר היחסי של הקרינה בתחום של 700-300 ננומטר ומיקמנו את המרכז של כל טיפוס צבע על 'משושה הצבעים של הדבורים' (Chittka 1992 ) כאשר צבע הרקע הממוקם במרכז המשושה הוא ירוק של עלים (איור 2). ככל שהמרחק (Euclidian distance) בין המרכזים של שני צבעים סמוכים גדול יותר, כך מסוגלות הדבורים להפריד יותר טוב ביניהם. סף ההפרדה לדבורים נע בין הערכים 0.062 (Dyer and Chittka 2004) לבין 0.100 (Chittka et al. 1997).
מספר השחלילים
פרח בודד של כלנית המצויה מכיל שחלילים (עלי שחלה מפורדים) רבים. על מנת להשוות את מספר השחלילים בפרחים מצבעים שונים ספרנו את השחלילים בחמישה פרחים מכל צבע (אדום לבן וכחול) ובדקנו בניתוח שונויות (ANOVA) האם היה הבדל מובהק.
מנגנון ההאבקה
האבקה עצמית ספונטנית
לבדיקה האם קיימת בכלנית האבקה עצמית ספונטנית השתמשנו ב 20 פרחים מכל צבע (אדום, לבן וכחול) בבית לחם הגלילית. פרחים צעירים כוסו ,טרם פתיחתם, ברשת עדינה מאוד (0.05X0.05 מ"מ) שמנעה כניסת חרקים וגרגרי אבקה ברוח. הפרחים כוסו ב 25.01.2010 וב 21.01.2011 ונפתחו לאחר 45 ימים לבדיקה של חנטת הפירות.
האבקת רוח
לבדיקה האם קיימת בכלנית האבקת רוח, הכנו 30 מלכודות אבקה מרצועות פלסטיק שבהם היו חורים עגולים, כל אחד בשטח של 28.27 ממ"ר עם סרט דביק מאחוריהם. המלכודות נתלו שלוש פעמים (16.01.2011, 26.02.2011, 6.03.2011) למשך 24 שעות, בין פרחי כלניות באוכלוסייה מעורבת מגידו. לאחר מכן, בבדיקה מיקרוסקופית זיהינו את גרגירי האבקה של כלנית מצויה על פי צורתם וגודלם, ספרנו אותם וחישבנו את קצב הגעתם לשעה לממ"ר.
לבדיקת שיעור האבקת רוח הצבנו באתר מגידו ב 2012 11כלובים (40X40X70 ס"מ) מכוסים ברשת (1X1 מ"מ), שמנעו כניסת חרקים אבל לא מנעו כניסת רוח שיכולה לשאת גרגרי אבקה. בבדיקה באתר זה לא נמצאו תריפסים העשויים להעביר גרגירי אבקה. בכל כלוב היו בממוצע (±ס.ת.) 3.2±1.95 פרחים מצבעים שונים. ב 2014 חזרנו על הניסוי בבית לחם הגלילית עם 12 צמחים אקראיים מכל צבע (אדום, לבן וכחול). שישה צמחים היו בכלובים למניעת כניסת חרקים ועוד שישה הושארו להאבקה חופשית. הניסוי הוצב ב 29.01.2014 וב 2.03.2014 נספרו מספר הפרחים שחנטו פירות (אך לא נספרו השחלילים בכל פרח). הניסוי נערך בחלקה של 100X200 מ' שהייתה בשדה אחיד חשוף לרוח.
ביקורי חרקים
תצפיות ראשונות לזיהוי החרקים המבקרים בפרחים ותדירות ביקורם בפרחים מצבעים שונים (אדום, לבן וכחול) בוצעו בפברואר ומרס 2011 ו 2012. שמונה עשרה תצפיות נוספות במשך כולל של 37 שעות בוצעו ב 2014 בשישה אתרים (טבלה 1). בכל אוכלוסייה בוצעו שלוש תצפיות בתאריכים שונים וכל תצפית נמשכה בממוצע ( ±ס.ת.) ±45124 דק'. על מנת לבדוק האם קיימת העדפה של חרקים מבקרים לפרחים בעלי צבע מסוים, עקבנו אחר סדר הביקורים של כל חרק מבקר, עד שעזב את שטח התצפיות (1,000 מ"ר). בכל סדרת תצפיות ספרנו בחלקה גם את מספר הפרחים מכל צבע (אדום, לבן או כחול) לאורך חמישה חתכים של 100 מ' כל אחד. התצפיות במבקרי הפרחים בוצעו רק בימי שמש נטולי רוח חזקה ובטמפרטורה גבוהה מ 20°C. השתמשנו במבחן פירסון (Pearson's χ2) לבדוק האם היה הבדל מובהק בין התפלגות התדירות של הצבעים השונים בשדה לבין התפלגות הביקורים של החרקים בהם (טבלה 1). אם אין הבדל מובהק פרושו שהחרק ביקר בפרחים בתדירות הדומה לתדירותם בשדה, כלומר ללא העדפה של צבע מסוים.
ההרכב הגנטי של האוכלוסיות ושל טיפוסי הצבע השונים
השונות הגנטית בין עשר האוכלוסיות שנדגמו (טבלה 1) ובין טיפוסי הצבע (אדום, לבן וכחול) נבדקה באמצעות ״טביעת אצבע גנטית״ בטכניקת Amplified Fragment Length Polymorphism (AFLP) . לביצוע השיטה עקבנו אחר הפרוטוקול המתואר אצל Vos et al. (1995). פירוט נוסף של כל שיטות העבודה ומקורות הספרות המדעית עליהם מתבססות הבדיקות הגנטיות והניתוח הסטטיסטי שלהן, ראו ב Dafni et al. (2020). בבדיקות גנטיות מהסוג הזה מקובל לדגום גם מין אחר קרוב על מנת לאמת שהשיטה אכן מבחינה בהבדלים. מכיוון שבישראל אין מין נוסף של כלנית, לא נכללה קבוצה כזו במחקר. אולם במחקרים קודמים שבוצעו באותה מעבדה ובאותה שיטה באורן ירושלים ובחלמונית גדולה התקבלו פרופילים גנטיים שונים לחלוטין. נוסף לכך, העובדה שבמחקר הנוכחי התגלתה שונות גנטית בין האוכלוסיות שנדגמו מהווה בקרה מספקת לאמינות השיטה.
תוצאות
החזר גלי האור מהפרחים וראייתם על-ידי דבורים
החזר גלי האור מפרחים אדומים, לבנים וכחולים של כלנית מצויה מאוכלוסיית מגידו היה שונה באופן בולט (איור 3) וכך גם המיקום שלהם על משושה ראית הצבעים של דבורי דבש (Chittka 1992) (איור 4). המיקום הממוצע (±ס.ת.) של פרחים אדומים (n=20) היה X=−0.08850±0.08869 Y=−0.03450±0.01050, של פרחים לבנים (n=20) X=0.08450±0.03069 Y=0.14350±0.02207 ושל פרחים כחולים (n=20) X=−0.1730±0.04438 Y=0.1675±0.03024. המרחק הממוצע (±ס.ת.) בין המיקום של פרחים אדומים וכחולים היה 0.261±0.050, בין לבנים וכחולים 0.287±0.050 ובין אדומים ללבנים היה 0.269±0.079.
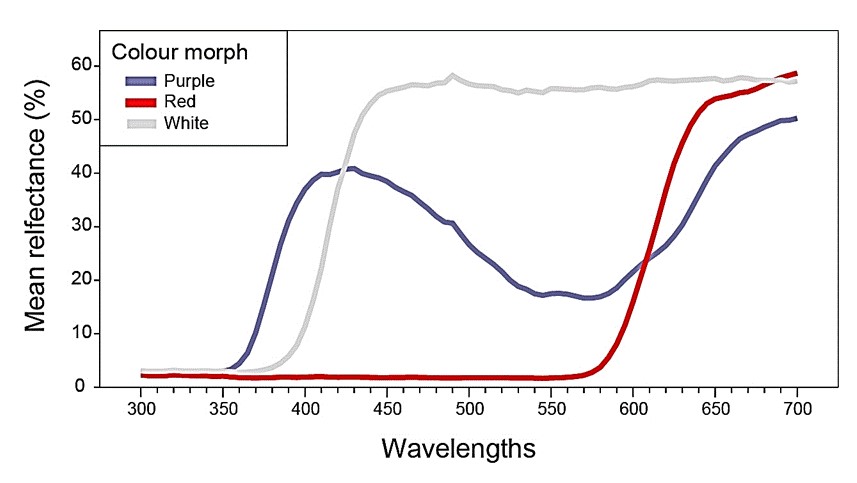
איור 3. החזר האור היחסי (%) מפרחים אדומים לבנים וכחולים מאתר מגידו.

איור 4. המיקום של פרחים אדומים, לבנים וכחולים של כלנית מצויה במשושה ראית הצבעים של דבורי דבש (Chittka 1992). מרכז המשושה מציין את הרקע של עלים ירוקים.
מספר השחלילים
מספר השחלילים (עלי שחלה מפורדים) הממוצע (±ס.ת.) של חמישה פרחים מכל צבע במגידו היה: אדום 584.6±75.7, לבן 582.1±69.1 וכחול 573.8±50.3. בניתוח שונויות (ANOVA) ההבדל לא היה הבדל מובהק (F2,14=0.007, P=0.993).
מנגנון ההאבקה
האבקה עצמית ספונטנית
בתנאי ניסוי בו נמנעו האבקת חרקים והאבקת רוח בבית לחם הגלילית ,לא הייתה כלל חנטת פירות ב 20 פרחים מכל צבע (אדום, לבן וכחול), כלומר לא הייתה האבקה עצמית ספונטנית בכלניות בכל טיפוסי הצבע.
האבקת רוח וחרקים
אחוז חנטת הפירות הממוצע (±ס.ת.) במגידו ב 2012 בכלובים (n=11) שמנעו גישת מאביקים היה 78±10. אחוז חנטת הפירות הממוצע בבית לחם הגלילית ב 2014 בהאבקה חופשית (חרקים ורוח) ב 46 פרחי כלניות, 18 מכל צבע היה בין 95.8% ל 100%. אחוז החנטה תחת כלובי רשת שמנעו האבקת חרקים אך אפשרו האבקת רוח ב 46 פרחי כלניות, 18 מכל צבע היה בין 90.3% ל 96.1%. לא היה הבדל מובהק בחנטת הפירות בין הכלניות מטיפוסי הצבע השונים בתנאים של האבקה חופשית (Kruskal–Wallis, χ=1.11, df=2, P=0.775) ולא נמצא הבדל גם בהאבקת רוח בלבד (Kruskal–Wallis, χ=1.72, df=2, P=0.632). לעומת זאת היה הבדל קטן בין אחוז חנטת הפירות הממוצע (±ס.ת.) בפרחים מכל הצבעים יחדיו, בתנאי האבקה חופשית (98.0±1.2) לעומת תאים של האבקת רוח בלבד (93.0±1.7). ההבדל הזה היה מובהק מבחינה סטטיסטית (Mann–Whitney U=160, Z=2,519, P=0.012, N=24).
בשלוש תקופות של 24 שעות, נלכדו בממוצע (±ס.ת.) 36.3±0.6 גרגירי אבקה של כלנית מצויה בכל מלכודת (30n=). הקצב המחושב של קליטת גרגירי האבקה במלכודת היה 0.05 גרגירים לשעה לממ"ר. שטח הפנים הממוצע (±ס.ת.), של צלקת בפרח של כלנית מצויה, כפי שחושב לאחר מדידה מיקרוסקופית היה 0.491±0.0092 ממ"ר. הצלקות של כלנית מצויה רצפטיביות במשך 3.5 ימים, כלומר כ 84 שעות (Dafni et al. 1990). מכך חישבנו שבמשך חיי פרח אחד של כלנית מצויה יכולה כל אחת מהצלקות, המכילה ביצית אחת, לקבל כ 2.1 גרגירי אבקה.
ביקורי חרקים בפרחים
בשנים 2011 ו 2012 מרבית המבקרים היו דבורי דבש, דבורים יחידאיות וזבובים, וכמעט שלא נצפו חיפושיות-פרחים Glaphyridae. בסך הכול נחתו רק 175 חרקים על פרחים מכל הצבעים, וקצב הביקורים הממוצע (±ס.ת.) היה 0.072±0.078 ביקורים לפרח לשעה. שיעור הביקורים הנמוך לא אפשר לנתח את הנתונים ולבדוק האם הייתה העדפה של פרחים מצבע מסוים על-ידי החרקים המבקרים.
ב 2014 חזרנו וערכנו ארבעה סבבים בלתי תלויים של תצפיות: שתי תצפיות ב 6.02.2014, תצפית אחת ב 9.02.2014 ועוד תצפית ב 21.02.2014. בביקורים אלה עקבנו אחר סדר הביקורים של 13, 120, 30 ו 43 חרקים בפרחי כלנית אדומים, לבנים וכחולים בהתאמה (טבלה 2). דבורי דבש, דבורים יחידאיות (מרביתן מהסוג Andrena) וזבוי רחף Syrphideae ביקרו בעיקר בפרחים לבנים ונמנעו גם הם מלבקר בפרחים אדומים, אולם המבחן הסטטיסטי היה רק כמעט מובהק. בעונה זו נצפתה רק חיפושית אחת מבקרת בפרח אדום (טבלה 2). על פי מבחן Spearman, דבורי דבש ודבורים יחידאיות .Andrena sp לא בקרו באקראי בפרחים בצבעים השונים בעיקר משום שהן נמנעו מלבקר בפרחים האדומים (טבלה 2).
טבלה 2. מספר הביקורים (נחיתות) של החרקים השונים בפרחים אדומים, לבנים וכחולים של כלנית מצויה ב 2014, סך כל מספר הפרחים מכל צבע בשלושת אתרי התצפית יחדיו, ותוצאות מובהקות המבחן (P value of Spearman's χ2 test with df=2) לבדיקת ההתאמה בין תדירות צבעי הפרחים ותדירות הביקורים בהם. 0.05>P מראה על הבדל מובהק כלומר על העדפה של צבע מסוים על-ידי החרק המבקר.
| מבקרים | אדום | לבן | כחול | ס"ה | P |
| דבורי דבש | 0 | 520 | 252 | 772 | <0.005 |
| Andrena sp. | 0 | 75 | 16 | 91 | <0.005 |
| זבובי רחף | 0 | 24 | 7 | 31 | 0.056 |
| חיפושיות פרחים | 1 | 0 | 0 | 1 | |
| ס"ה ביקורים | 1 | 619 | 275 | 895 | |
| מספר הפרחים | 115 | 694 | 377 | 1015 |
ההרכב הגנטי של האוכלוסיות ושל טיפוסי הצבע השונים
האנליזה של ה DNA בשיטת AFLP חשפה 462 אתרים פולימורפיים בגנום של כלנית מצויה. רמת הפולימורפיזם (P) והשונות הגנטית (He-unbiased expected heterozygosity) של כל טיפוס צבע בכל אוכלוסייה מוצגים בטבלה 3. אחוז הפולימורפיזם המומצע (±ש.ת.) היה 70.67±5.25=P והשונות הגנטית הייתה He=0.124±0.002. מידת הזהות הגנטית (I – Nei unbiased genetic identity) בין האוכלוסיות הנבדקות: הר-מירון, צפורי, אוניברסיטת חיפה, דרך נוף כרמל, אלונים, בית שערים, בלפוריה, מגידו, פורה ולהב הייתה גבוהה מאוד (I=0.99) המלמדת על העדר הבדלים בין האוכלוסיות ולא בין טיפוסי הצבעים השונים (Exact test, P>0.99).
אנליזה של השונות הגנטית (AMOVA) (Peakall and Smouse, 2006 ) הראתה ש 99% מהשונות הגנטית מקורה בשונות בתוך האוכלוסיות ורק 1% מקורו בשונות בין האוכלוסיות (F9,226=0.014, P=0.001). המבחן הסטטיסטי Bayesian assignment test (Pritchard et al. 2000; Falush et al. 2007) הראה גם הוא העדר הבדלים בין האוכלוסיות ולא בין טיפוסי הצבע השונים. מבחן זה מחלק את כלל הנתונים למספר קבוצות (K) והחלוקה המיטבית (הנקבעת על פי הערך של Delta K) הייתה לשתי קבוצות (33.8) או ארבע קבוצות (32.6). גרף המתאר את ההרכב הגנטי של כל אתר בחלוקה למספר שרירותי של 2, 3 ו 4 קבוצות (STRUCTURE Bayesian clustering) מראה דמיון גדול בגרף העמודות בין כל הפרטים בכל האוכלוסיות (איור 5).
טבלה 3. מספר הצמחים בדגימה (N), השונות הגנטית הממוצעת (He=unbiased expected heterozygosity), שגיאת התקן ואחוז הפולימורפיזם (P) בכל האוכלוסיה (מצפון לדרום), בטיפוסים בעלי פרחים כחולים, לבנים או אדומים, ועבור בכל הצבעים יחדיו על פי אנליזת AFLP של 462 אתרים פולימורפיים.
| כחול | לבן | אדום | כל הצבעים | ||||||||||
| האתר | SE | He | N | SE | He | N | SE | He | N | P(%) | SE | He | N |
| הר מירון | 0.008 | 0.191 | 8 | 0.008 | 0.133 | 6 | 0.007 | 0.139 | 14 | 87.7 | 0.007 | 0.156 | 28 |
| ציפורי | 0.007 | 0.114 | 15 | 0.007 | 0.153 | 14 | 0.006 | 0.109 | 12 | 89.2 | 0.007 | 0.141 | 41 |
| אוניברסיטת חיפה | — | — | — | — | — | — | 0.006 | 0.115 | 15 | 66.9 | 0.006 | 0.115 | 15 |
| דרך נוף כרמל | 0.007 | 0.153 | 11 | — | — | — | 0.007 | 0.096 | 8 | 77.9 | 0.007 | 0.136 | 19 |
| אלונים | 0.008 | 0.154 | 5 | 0.009 | 0.109 | 3 | 0.007 | 0.096 | 11 | 67.8 | 0.006 | 0.12 | 19 |
| בית שערים | — | — | — | 0.01 | 0.097 | 2 | 0.007 | 0.14 | 10 | 63.6 | 0.007 | 0.14 | 12 |
| בלפוריה | 0.008 | 0.101 | 8 | 0.007 | 0.094 | 12 | 0.005 | 0.074 | 15 | 60.6 | 0.006 | 0.094 | 35 |
| מגידו | 0.007 | 0.178 | 12 | 0.008 | 0.177 | 11 | 0.006 | 0.077 | 11 | 93.7 | 0.006 | 0.15 | 34 |
| פורה | — | — | — | — | — | — | 0.007 | 0.097 | 14 | 60 | 0.007 | 0.108 | 18 |
| להב | — | — | — | — | — | — | 0.007 | 0.085 | 14 | 39.4 | 0.007 | 0.084 | 15 |
| Total | 0.007 | 0.146 | 59 | 0.007 | 0.136 | 48 | 0.006 | 0.102 | 124 | 70.7 | 0.002 | 0.124 | 236 |
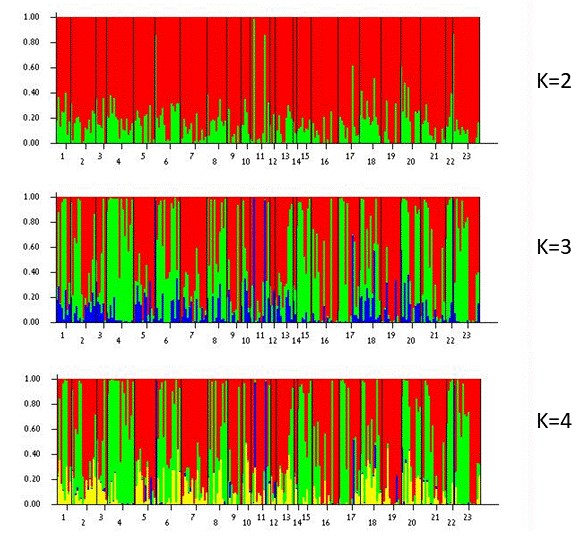
איור 5. עקומת עמודות (STRUCTURE Bayesian clustering), המקבצת את 462 האתרים הפולימורפיים בכלנית המצוייה לשתיים (K=2), שלוש (K=3) או ארבע (K=4) קבוצות. המספרים בציר האופקי מציינים כל אחד טיפוס צבע באוכלוסיה. 1 – הר מירון אדום, 2 – הר מירון לבן, 3 – הר מירון כחול, 4 – ציפורי אדום, 5 – ציפורי לבן, 6 – ציפורי כחול, 7 – אוניברסיטת חיפה אדום, 8 – דרך נוף כרמל אדום, 9 – דרך נוף כרמל כחול, 10 – אלונים אדום, 11 – אלונים לבן, 12 – אלונים כחול, 13 – בית שערים אדום, 14 – בית שערים לבן, 15 – בלפוריה אדום, 16 – בלפוריה לבן, 17 – בלפוריה כחול, 18 – מגידו אדום, 19 – מגידו לבן, 20 – מגידו כחול, 21 – פורה אדום, 22 – פורה אדום-כתום, 23 – להב אדום.
דיון
ההבחנה בין צבעי הכלניות על-ידי החרקים
לכלנית המצויה בישראל טיפוסי צבע שונים (איור 1) הנבדלים באופן ברור לעין האנושית ובעקומת ההחזר היחסי של האור באורכי גל שונים (איור 2). למרות שלעין האדם צבעי הכלניות נראים שונים, אין זה אומר שגם דבורים מבחינות בצבעים אלה. זאת משום שהרגישות של קולטני האור (פוטורצפטורים) בעיניים של בעלי חיים שונים, לאור באורכי גל שונים אינה דומה. כך למשל עין האדם רגישה לצבע אדום אך לא לאולטרה סגול ועין הדבורה רואה אולטרה סגול אך לא אדום. לכן, על מנת לבדוק האם בעל חיים מסוים מבחין בין צבעים שונים יש להשתמש במודלים הלוקחים בחשבון את הרגישות היחסית של הקולטנים שלו לאור באורכי גל שונים (Chittka 1992). ככל שהמרחק בין שני צבעים גדול יותר כך היכולת להבחין בין הצבעים טובה יותר וקיים ערך סף שמתחתיו כנראה שבעל החיים מתקשה, או אינו יכול להבחין בהבדל בין הצבעים. לגבי דבורים נמצא שערך סף זה נע בין 0.062 (Dyer and Chitka 2004) לבין 0.100 (Chittka et al. 1997). המרחק, במשושה הצבעים של דבורי דבש, בין כלניות אדומות לבנות וכחולות היה בין 0.26 ל 0.27, כלומר דבורי דבש וכנראה גם דבורים יחידאיות מסוגלות להבחין היטב ביניהן. מרכז המשושה מייצג את צבע הרקע של עלים ירוקים וככל שהצבע ממוקם רחוק יותר ממנו כך הוא גם בולט יותר לעין הדבורה. מכיוון שמיקום הצבע האדום של הכלניות האדומות קרוב יותר למרכז, זה אומר שדבורים מבחינות בהן פחות טוב מאשר כלניות לבנות או כחולות.
בניגוד לדבורים, נתונים על ראיית צבע בזבובים ויכולת ההבחנה שלהם בין צבעים שונים (ללא קשר לבהירותם) נחקרה רק מעט (Lunau 2014). לכן לא יכולנו לבדוק באיזו מידה יכולים זבובי הרחף Syrphidae שביקרו בפרחי הכלניות להבחין בין טיפוסי הצבע השונים.
חיפושיות-הפרחים Glaphyridae לא נמצאו מבקרות בפרחי כלניות במחקר הנוכחי שנערך בתחילת העונה בה מופיעות בעיקר הכלניות הכחולות והלבנות, לפני שיא פריחת הכלניות האדומות ופעילות החיפושיות המקבילה להן. במחקרים אחרים נמצא קשר הדוק בין החיפושיות לבין הפרחים האדומים המספקים להם מזון, מסתור ומקום הזדווגות (Dafni et al. 1990; Keasar et al. 2008). לחיפשיות פרחים אלה יש קולטנים הרגישים לאדום, ירוק ואולטרה סגול (Martinez-Harms et al. 2012).
ההבדלים באור המוחזר מפרחי כלניות בצבעים שונים, וההבדלים ברגישות של החרקים השונים (דבורים זבובים וחיפושיות) לאור באורכי גל שונים, מעידים על הבדלים גם ביכולתם לגלות ולהבדיל בין הפרחים בצבעים השונים. הבדל זה יכול היה להיות בסיס פיסיולוגי להעדפות השונות שלהם לבקר בפרחי כלנית מצבעים שונים, ובכך להסביר את קיום טיפוסי הצבע השונים זה לצד זה באותם בתי גידול.
האבקה עצמית ספונטנית והאבקת רוח
במחקר קודם (Horovitz 1995) נמצאה אי סבילות (עקרות) עצמית בכלנית המצויה כלומר אי-יכולתה לייצר זרעים כתוצאה מהאבקה עצמית. כמו כן נמצא שהאבקה בין צמחים מטיפוסי צבע שונים היא פורייה ומייצרת אגוזיות בעלות זרעים חיוניים. תוצאות המחקר הנוכחי הראו גם הם העדר יצור זרעים בהאבקה עצמית ספונטנית בכלנית מצויה. אבל, לעומת זאת, הייתה חנטה כמעט מלאה (98%) בפרחים שהיו נתונים להאבקה חופשית של חרקים ורוח וחנטה נמוכה רק במעט (93%) בפרחים שהיו נתונים להאבקת רוח בלבד וללא האבקת חרקים. תוצאות אלה הפתיעו אותנו, משום שפרחי הראווה של הכלנית משייכים אותה לסינדרום של פרחים המאבקים על-ידי חרקים ולא על-יד רוח (Horovitz 1990). בתנאים אלה ייתכן, שלמרות הצבעים השונים והתנהגות הדיפרנציאלית של המאביקים, האבקת רוח מאפשרת חילוף גנים חופשי בין טיפוסי הצבע השונים של הכלנית המצויה. מכיוון שהכלנית היא צמח נפוץ בעל הפצת זרעים על-ידי הרוח, האבקת רוח עשויה לתרום למיסוך ההשפעה של התנהגות מאביקים שעשויה הייתה לגרום לבידוד רבייתי וגנטי בין טיפוסי הצבע בכלנית.
למרות הניגוד הגדול בין התכונות של פרחים מאבקי חרקים (גדולים, צבעוניים וגומלים, בעלי גרגירי אבקה גדולים ודביקים) לתכונות של מאבקי רוח (קטנים, ירקרקים ובלתי גומלים, בעלי גרגירי אבקה קטנים, חלקים ושאינם דביקים) יש צמחים בהם נמצאה האבקת רוח בצירוף עם האבקת חרקים, תופעה המכונה 'אמבופיליה' (Ambophily) (Regal 1982; Dafni et al. 2012). האמבופיליה מיוחסת לתנאים סביבתיים בהם יש קשיים לפעילות של חרקים מאביקים. תכונה זו התפתחה באופן בלתי תלוי לפחות שלוש פעמים בסוגים שונים במשפחת הנוריתיים אליה שייכת גם הכלנית המצויה (Kaplan and Mulcahy 1971; De la Bandera and Traveset, 2006; Duan et al. 2009). החורף הים-תיכוני הוא מתון, לכן קשה להסביר את האבקת הרוח בכלנית כהתאמה לתנאי סביבה קשים. ייתכן שהאבקת רוח בכלנית המצויה התפתחה כמנגנון 'ביטוח' למקרה של תנאים גשומים המצמצמים מאוד פעילות של מאביקים (Dafni 1996 ). פרחי הכלנית נסגרים בלילה ובימים גשומים, וכך נשמרת האבקה יבשה, והאבקת רוח יכולה להתקיים בימי שמש יבשים שאינם נדירים בחורף הים-תיכוני.
ביקורי חרקים בפרחים
הכלניות האדומות נחקרו בעבר, וחיפושיות הפרחים (Glaphyridae) נמצאו כמאביק העיקרי שלהן (Dafni et al. 1990). למרות זאת בתחילת העונה, כשמקורות האבקה במיני פרחים אחרים דלים, נמצא שכלניות האדומות מבוקרות גם על-ידי דבורי דבש (Keasar et al. 2008). במחקר הנוכחי דבורי הדבש היו המבקרות העיקריות בפרחי הכלנית, לאחריהן דבורים יחידאיות בעיקר מהסוג Andrena sp., זבובי רחף, ורק חיפושית אחת נמצאה בפרחים (טבלה 2). ההבדלים הבולטים בהרכב חברת החרקים המבקרים ובתדירות הביקורים של המינים השונים במחקר הנוכחי ובמחקרים קודמים (Keasar et al. 2008; Keasar et al. 2010; Keasar et al. 2013 ) עשויים לשקף את השונות הגדולה בטמפרטורות, במשקעים ובמינים נוספים הפורחים בו זמנית יחד עם הכלניות בחורף הים-תיכוני.
ההרכב הגנטי של האוכלוסיות ושל טיפוסי הצבע השונים
האנליזה הגנטית של עשר אוכלוסיות של כלנית מצויה הכוללות צמחים בעלי פרחים אדומים, לבנים וכחולים הגדלות לאורך מפל הגשם בישראל חשפה רמה גבוה של פולימורפיזם ושונות גנטית (טבלה 3). במקביל נמצא דמיון גנטי גבוה בין האוכלוסיות השונות ובין טיפוסי הצבע (אדום לבן וכחול). ניתוח הנתונים הראה שכמעט כל השונות הגנטית (99%) מקורה בשונות בתוך האוכלוסיות, ולא נמצאו הבדלים מובהקים בהרכב הגנטי בין אוכלוסיות ולא בין טיפוסי הצבע. המסקנה מתוצאות אלה היא שבכלנית המצויה בישראל קיימת זרימה חופשית של גנים בין האוכלוסיות ובין טיפוסי הצבע. מסקנה זו מעלה את השאלה מה הם הגורמים האבולוציוניים והמנגנונים האקולוגיים שהביאו להתפתחות הפולימורפיות הקיצונית בצבע הפרחים בכלנית המצויה ומשמרים אותה.
האבולוציה והאחזקה של פולימורפיות בצבעי פרחים
המנגנונים העשויים לגרום להתפתחות של פולימורפיזם קיצונית בצבע הפרחים בתוך או בין אוכלוסיות של צמחים מאותו מין הם מגוונים. כך למשל שינויים מרחביים או זמניים בעוצמה ובכיוון של הברירה הטבעית על צבע הפרחים עשויה לתחזק שונות גנטית כזו (Hedrick et al. 1976; Hedrick 1986; Ellner and Sasaki 1996 ). סלקציה המופעלת על-ידי מאביקים עשויה להסביר פולימורפיזם בצבע פרחים בכך שלמאביקים שונים יש העדפות מולדות או נלמדות לפרחים בעלי צבעים שונים (7,11,21,30,80). מאביקים המבחינים ומעדיפים, באופן מולד או נלמד, לבקר בטיפוסי צבע שונים במסגרת של אותו מין צמח, הם עשויים לגרום בהתנהגותם לסלקציה ולהפרדה בין טיפוסי הצבע השונים, כפי שהודגם לגבי דבורים פרפרים וציפורים (Kay 1976; Stanton 1987; Irwin and Strauss 2005; Raine and Chittka 2007). רבגוניות בצבע הפרחים עשויה להישמר גם במקרים בהם לפרטים שונים של מין מסוים של מאביק, הפעילים במקומות או זמנים שונים, יש העדפות שונות לפרחים בעלי צבעים שונים. כך יכולה לטיפוס צבע מסוים להיות כשירות גבוהה יותר בעונה או מקום מסוים.
לעיתים, צבע הפרחים במין מסוים עשוי להיות קשור בתאחיזה לתכונות אחרות המשפיעות ישירות על כשירות הצמח. במקרים כאלה, צבע הפרחים יכול להיקבע בעיקר על-ידי התכונות הקשורות אליו יותר מאשר על-ידי ברירה טבעית ישירה על צבע הפרח עצמו (Fineblum and Rausher 1997; Armbruster 2002; Strauss and Whittall 2006).
תוצאות מחקר זה מביאות עדות ברורה להעדפה דיפרנציאלית של טיפוסי צבע שונים על-ידי המאביקים. דבורי דבש ודבורים יחידאיות מהסוג Andrena נמנעו מלבקר בפרחים האדומים (טבלה 1), זאת בניגוד לממצאים קודמים (Keasar et al. 2008), אבל הם ביקרו בפרחים לבנים וכחולים על פי המצופה ותדירותם. ההבדל בהתנהגות דבורי הדבש המשתקף בתוצאות המחקרים השונים עשויי להיות מוסבר בהבדלים בתנאי הסביבה, או גם בהבדלים בהרכב במינים האחרים שפרחו במקביל ובשפע שלהם. לכן, ההתנהגות וההאבקה על-ידי דבורי הדבש יכולה לתרום רק באופן שולי לבידוד טיפוסי הצבע השונים בכלנית. זבובי הרחף נמנעו גם הם מלבקר בפרחים האדומים אולם ללא העדפה מובהקת מבחינה סטטיסטית לצבע מסוים (טבלה 2). חיפושיות הפרחים (Glaphyridae), שנעדרו כמעט לחלוטין מהחרקים מבקרי הפרחים במחקר זה, נמצאו במחקרים קודמים כמבקרים עיקריים של פרחי כלניות אדומות (Dafni et al. 1990). גם בהעדר העדפה ברמה של מין החרק המבקר, הנאמנות של דבורי דבש למקור מזון גומל במשך מעוף לחיפוש מזון עשויה לגרום לזרימת גנים רבה יותר בין צמחים בעלי אותו צבע פרחים מאשר בין צמחים בעלי פרחים בצבע שונה. כמו כן, מכיוון שההעדפה של מאביקים תלויה גם בתדירות הפרחים, העדפה זו עשויה להשתנות בין מקומות שונים, עונות או שנים שונות. כלומר, על פניו נראה שלפעילות המבקרים בפרחים עשויה הייתה להיות תרומה לאבולוציה של הפולימורפיזם בצבע פרחי הכלניות ולשימורה. אבל כפי שצוין קודם, האבקת הרוח פועלת לזרימת גנים חופשית בין טיפוסי הצבע ובין האוכלוסיות.
הכלנית המצויה פורחת בחורף, בזמן שפעילות המאביקים נמצאת בשפל (Dafni 1996). במקביל לשפע הכלניות האדומות יש גם שפע של חיפושיות הפרחים (Glaphyridae) המאביקות אותן (Dafni et al. 1990). אבל במקביל, בעיקר בשל דלות פרחי אבקה אחרים דבורי הדבש אוספות אבקה בפרחי הכלניות האדומות (Keasar et al. 2008 ). לו היו חיפושיות הפרחים נפוצות רק בדרום ישראל, זה היה עשוי להסביר את מציאות האוכלוסיות בעלות הפרחים האדומים בדרום בלבד. אבל החיפושיות נפוצות גם בצפון ישראל מקומות בהם יש כלניות במגוון צבעים (דפני מידע אישי).
העובדה שהאוכלוסיות הדרומיות של הכלנית המצויה בישראל הן אדומות בלבד העלתה את ההשערה שהצבע האדום קשור לעמידות הצמחים ליובש (Horovitz et al. 1975; Horovitz 1976). רמז אפשרי לכך הוא שלצמחי הכלניות האדומות בדרום יש עלים צרים יותר מאשר לאלה הגדלות בצפון (טנקוס מידע אישי), למרות שחסר מידע האם זו תכונה תורשתית או ביטוי של פלסטיות פנוטיפית.
העמידות של הכלניות האדומות לרעייה כבדה של בקר (Perevolotsky et al. 2011) עשויה להוות יתרון על פני טיפוסי צבע אחרים. אולם משום שבית הגידול של הכלנית המצויה הוא בשטחים עשבוניים פתוחים, שמרביתם נתונים לרעייה מסיבית, עמידות זו לא יכולה להסביר למה חלק מהאוכלוסיות הגדלות בצפון הן מגוונות וחלקן אדומות בלבד. בהקשר לרעייה, נמצא שאייל הכרמל, בניגוד לבקר, אוכל פרחים של כלניות אדומות.(Wallach et al. 2009)
התפוצה של טיפוסי הצבע השונים של הכלנית המצויה בישראל קשורה כנראה גם לסוג הסלע והקרקע. אוכלוסיות בעלות פרחים לבנים בלבד גדלות על קרקעות נטולות גיר חופשי על גבי סלעי גיר קרסטי (תצורת בר-כוכבא) באזור עמיעד-ראש פינה. במקביל, הכלניות בגולן, הגדלות גם הן על גבי קרקעות בזלתיות דלות בגיר, הן בעיקר בעלות פרחים בגוונים שונים של כחול-סגול-ארגמן (Horovitz et al. 1975; Horovitz 1976; Horovitz 1990). אבל בגליל ובכרמל קיימות אוכלוסיות כלניות אדומות בלבד הגדלות לעיתים בסמיכות לאוכלוסיות מגוונות על גבי סלעים וקרקעות דומים. לכן נראה שנושא זה דורש מחקר יסודי נוסף.
לסיכום, אין לנו הסבר להתפתחות האבולוציונית של רבגוניות קיצונית בצבעי הפרחים של הכלניות ולא לדגם התפוצה שלהם בישראל. למה בדרום גדלות אוכלוסיות של כלניות אדומות בלבד, בעוד בצפון גדלות אוכלוסיות אדומות לצד מגוונות. גם החוקרים Narbona et al. (2017), שסקרו את נושא הפולימורפיות בצבעי פרחים באגן הים-התיכון (אבל משום מה לא הזכירו את הכלנית המצויה) אינם מציעים מנגנון להתפתחות התופעה אלא מציינים ש "העובדה שלא נמצא מקרה של מין רק עם אוכלוסיות מגוונות, מרמז על כך שצבע הפרחים במינים פולימורפיים נתון לתהליכים אבולוציוניים שונים בתחום תפוצתם".
תודות
מחקר זה נתמך על-ידי קרן הנק ודורותי שוסהיים למחקר אקולוגי בכרמל.
ספרות:
רשימת ספרות מפורטת נמצאת במאמר המקורי: Dafni et al. 2020
Arista M Talavera M Berjano R and Ortiz PL 2013 Abiotic factors may explain the geographical distribution of flower colour morphs and the maintenance of colour polymorphism in the scarlet pimpernel. Journal of. Ecology 101: 1613-1622.
Armbruster WS 2002 Can indirect selection and genetic context contribute to trait diversification? A transition-probability study of blossom-colour evolution in two genera. J. of Evolutionary Biology 15: 468-486.
Brown BA and Clegg MT 1984 The influence of flower color polymorphisms on genetic transmission in a natural population of the common morning glory, Ipomoea purpurea. Evolution 38: 796–803.
Chittka L 1992 The colour hexagon: a chromaticity diagram based on photoreceptor excitations as a generalized representation of colour opponency. Journal of Comparative Physiology A 170: 533-543.
Chittka L Gumbert A and Kunze J 1997 Foraging dynamics of bumble bees, correlates of movements within and between plant species. Behavioral Ecology 8: 239-249.
Chittka L Thomson JD and Waser NM 1999 Flower constancy, insect psychology, and plant evolution. Naturwissenschaften 86: 361-377.
Chittka L and Kevan PG 2005 Flower colour as advertisement. In: Dafni A. Kevan PG. Husband BC. Editors) Practical Pollination Biology. Enviroquest Ltd.: Cambridge, ON, Canada, pp. 157-196.
Dafni A Bernhardt P Shmida A et al. 1990 Red bowl-shaped flowers, convergence for beetle pollination in the Mediterranean region. Israel Journal of Botany 39: 81-92.
Dafni A 1996 Autumnal and winter pollination adaptations under Mediterranean conditions. Bocconea 5: 171-181.
Dafni A Marom-Levy T Jürgens A et al. 2012 Ambophily and “super generalism” in Ceratonia siliqua pollination. In: Patiny S. (editor) Evolution of Plant–Pollinator Relationships; Cambridge University Press, Cambridge, UK. pp: 344–373.
Dafni A Tzohari H Ben-Shlomo R Vereecken N and Ne’eman G 2020 Flower colour polymorphism, pollination modes, breeding system and gene flow in Anemone coronaria. Plants 9: 397. https://www.mdpi.com/2223-7747/9/3/397
De la Bandera MC and Traveset A 2006 Breeding system and spatial variation in the pollination biology of the heterocarpic Thymelaea velutina (Thymelaeaceae). Plant Systics and Evolution 257: 9-23.
Duan YW Zhang TF He YP and Liu JQ 2009 Insect and wind pollination of an alpine biennial Aconitum gymnandrum (Ranunculaceae). Plant Biology 11: 796-802.
Dyer AG and Chittka L 2004 Biological significance of distinguishing between similar colours in spectrally variable illumination, bumblebees (Bombus terrestris) as a case study. Journal of Comparative Physiology A 190: 105-114.
Dyer AG Spaethe J and Prack S 2008 Comparative psychophysics of bumblebee and honeybee colour discrimination and object detection. Journal of Comparative Physiology A 194: 617-627.
Ellner S and Sasaki A 1996 Patterns of genetic polymorphism maintained by fluctuating selection with overlapping generations. Theoretical Population Biology 50: 3-65.
Falush D Stephens M and Pritchard JK 2007 Inference of population structure using multilocus genotype data, dominant markers and null alleles. Molecular Ecology Notes 7: 574-578.
Fineblum WL and Rausher MD 1997 Do floral pigmentation genes also influence resistance to enemies? The W locus in Ipomoea purpurea. Ecology 78: 1646-1654.
Frey FM 2004 Opposing natural selection from herbivores and pathogens may maintain floral-color variation in Claytonia virginica (Portulacaceae). Evolution 58: 2426-2437.
Gegear RJ and Laverty TM 2001 The effects of variation among floral traits on the flower constancy of pollinators. In: Chittka L and Thomson JD (eds.) Cognitive Ecology of Pollination. Cambridge University Press: Cambridge, UK. pp 297-317.
Harvey DM 1971 Phenotypic variation in flower colour within the Anemone coronaria cultivars. Annals of. Botany 35: 1-8.
Hedrick PW 1986 Genetic polymorphism in heterogeneous environments, a decade later. Annual Review of Ecology and Systematics 17: 53–566.
Hedrick PW Ginevan ME and Ewing EP 1976 Genetic polymorphism in heterogeneous environments. Annual Review of Ecology and Systematics 7: 1-32.
Horovitz A 1976 Edaphic factors and flower colour distribution in the Anemone coronaria (Ranunculaceae). Plant Systematics and Evolution 126: 239-242.
Horovitz A 1990 The pollination syndrome of Anemone coronaria L. an insect-biased mutualism. VI Internationak Symposium on Pollination 288: 283-287.
Horovitz A 1995 The local gene pool of Anemone coronria. Ornamental Plant Improvement: Classical and Molecular 420: 144-146.
Horovitz A and Zohary D 1966 Spontaneous variegation for perianth colour in wild Anemone coronaria. Heredity 21: 513-515.
Horovitz A Galil J and Zohary D 1975. Biological flora of Israel, 6. Anemone coronaria L. Israel Journal of Botany 24: 26-41.
Irwin RE and Strauss SY 2005 Flower color microevolution in wild radish, evolutionary response to pollinator-mediated selection. American Naturalist 165: 225-237.
Johnson ET Berhow MA and Dowd PF 2008 Colored and white sectors from star-patterned petunia flowers display differential resistance to corn earworm and cabbage looper larvae. Journal of Chemical Ecology 34: 757-765.
Jones KN and Reithel JS 2001 Pollinator-mediated selection on a flower color polymorphism in experimental populations of Antirrhinum (Scrophulariaceae). Americam Journal of Botany 88: 447-454.
Joseph N and Siril EA 2013 Floral color polymorphism and reproductive success in annatto (Bixa orellana L.). Tropical Plant Biology 6: 217-227.
Kaplan SM and Mulcahy DL 1971 Mode of pollination and floral sexuality in Thalictrum. Evolution 25: 659-668.
Kay QON 1976 Preferential pollination of yellow-flowered morphs of Raphanus raphanistrum by Pieris and Eristalis spp. Nature 261: 230-232.
Keasar T Shmida A and Zylbertal A 2008 Pollination ecology of the red Anemone coronaria: honeybees may select for early flowering. Discussion Paper # 491. Center for the Study of Rationality: The Hebrew University of Jerusalem, Israel. https://www.researchgate.net/profile/Avi_Shmida/publication/23531070.
Keasar T Harari AR Sabatinelli G et al. 2010 Red anemone guild flowers as focal places for mating and feeding by Levant glaphyrid beetles. Biological Journal of the Linnaean Society 99: 808-817.
Keasar T Kishinevsky M Shmida A Gerchman Y et al. 2013 Plant-derived visual signals may protect beetle herbivores from bird predators. Behav Ecol Sociobiol , 67: 1613–1622.
Lunau K 2014 Visual ecology of flies with particular reference to colour vision and colour preferences. Journal of Comparative Physiology A 20: 497-512.
Malerba R and Nattero J 2012 Pollinator response to flower color polymorphism and floral display in a plant with a single-locus floral color polymorphism, consequences for plant reproduction. Ecological Research 27: 377-385.
Martinez-Harms J Vorobyev M Schorn J Shmida A et al. 2012 Evidence of red sensitive photoreceptors in Pygopleurus israelitus (Glaphyridae, Coleoptera) and its implications for beetle pollination in the southeast Mediterranean. Journal of Comparative Physiology A 198: 451-463.
Narbona E Wang H Ortiz PL et al. 2017 Flower colour polymorphism in the Mediterranean Basin, occurrence, maintenance and implications for speciation. Plant Biology 20: 8-20.
Peakall R and Smouse PE 2006 GENALEX 6, genetic analysis in excel. Population genetic software for teaching and research. Molecular Ecology Notes 6: 288-295.
Peitsch D Fietz A Hertel H de Souza J et al. 1992 The spectral input systems of hymenopteran insects and their receptor-based colour vision. Journal of Comparative Physiology A 170: 23–40.
Perevolotski AR Schwartz-Tzachor E Yonathan R and Ne’eman G 2011 Geophytes-herbivore interactions: reproduction and population dynamics of Anemone coronaria L. Plant Ecology 212: 563-571.
Pritchard JK Stephens M and Donnelly P 2000 Inference of population structure using multilocus genotype data. Genetics 155: 945-959.
Raine NE and Chittka L 2007 The adaptive significance of sensory bias in a foraging context, floral colour preferences in the bumblebee Bombus terrestris. PLoS ONE 2, e556. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0000556
Regal PJ 1982 Pollination by wind and animals: Ecology of geographic patterns. Annual Review of Ecology and Systematics 13: 497-524.
Schemske DW and Bierzychudek P 2007 Spatial differentiation for flower color in the desert annual Linanthus parryae, was right? Evolution 61: 2528-2543.
Stanton ML 1987 Reproductive biology of petal color variants of wild populations of Raphanus sativus L. I. pollinator response to color morphs. American journal of Botany 74: 176-185.
Steyn WJ Wand SJE Holcroft DM and Jacobs G 2002 Anthocyanins in vegetative tissues, a proposed unified function in photoprotection. New Phytologist 155: 349-361.
Strauss SY Irwin RE and Lambrix VM 2004 Optimal defence theory and flower petal colour predict variation in the secondary chemistry of wild radish. Journal of Ecology 92: 132–141.
Strauss SY and Whittall JB 2006 Non-pollinating agents of selection on floral traits. In: Harder LD. Barrett SCH. (editors) Ecology and Evolution of Flowers. Oxford University Press, Oxford, UK. pp. 120-138.
Vos P Hogers R Bleeker M et al. 1995 AFLP, a new technique for DNA fingerprinting. Nucleic Acids Research 3: 4407-4414.
Wallach AD Inbar M and Shanas U 2009 Roe deer and decapitated Anemone flowers. Israel Journal of Plant Sciences 2009, 57, 103–106.
Wang H Talavera M Min Y et al. 2016 Neutral processes contribute to patterns of spatial variation for flower colour in the Mediterranean Iris lutescens (Iridaceae). Annals of Botany 117: 995-1007.
=====================================
כל הזכויות שמורות ל"כלנית" ©
לציטוט: דפני א צהרי ח בן-שלמה ר ווריקן נ ונאמן ג 2020 צבע הפרחים, ההאבקה, והגנטיקה של הכלנית המצויה Anemone coronaria בישראל, כתב-עת "כלנית", מספר 7.










