גידי נאמן החוג לביולוגיה וסביבה, אוניברסיטת חיפה-אורנים, gneeman@gmail.com
תקציר: אלון התבור מוגדר כמין העומד בשלכת חורף, אולם קיימים הבדלים גדולים במועדי השרת העלים בין עצים סמוכים, בין אוכלוסיות ובין אותם עצים בין שנים. מעקב אחר הפנולוגיה של מועדי השרת העלים והלבלוב של כ 240 עצים בארבע אוכלוסיות במשך ארבע שנים חשף שההבדלים הללו נובעים כנראה מהבדלים גנטיים בין הפרטים וגם מהשפעה הפנוטיפית של תנאי הסביבה המשתנים משנה לשנה. לאור התוצאות ניתן לשער כי אלון התבור בישראל נמצא כנראה בשלב מעבר אבולוציוני ממין העומד בשלכת חורף למין ירוק עד. מאמר עברי זה מתבסס על מחקר מדעי שפורסם בעבר באנגלית:
Ne’eman 1993. Variation in leaf phenology and habit in Quercus ithaburensis, a Mediterranean deciduous tree. Journal of Ecology 81: 627-634
================= =
תקציר
במחקר זה עקבתי אחר הפנולוגיה (ההתנהגות העונתית) של מועדי השרת העלים בסתיו והלבלוב האביבי של כ 240 עצי אלון התבור Quercus ithaburensis בארבע אוכלוסיות במשך ארבע שנים. נמצאה שונות גדולה במשתנים הפנולוגיים (מועדי תחילה וסיום השרת העלים, משך השלכת, מועדי תחילה, סיום ומשך הלבלוב האביבי) בין העצים בתוך כל אחת מהאוכלוסיות. חלק מהעצים הקדימו להשיר את עליהם והיו בשלכת חורף במשך תקופה ארוכה יחסית לעצים אחרים אשר אחרו להשיר את העלים והייתה להם שלכת חורף קצרה בלבד. כ18% אחוז מהעצים סיימו להשיר את עליהם רק לאחר תחילת הלבלוב האביבי ולכן יכולים להיחשב כירוקי עד. נמצאו הבדלים מובהקים בערכים הממוצעים של משתני ההתנהגות העונתית של האוכלוסיות השונות וגם בין השנים בכל אחת מהן. נמצא מתאם בין המשתנים הפנולוגיים הממוצעים בכל אחת מהאוכלוסיות בשנים השונות לבין משתנים אקלים שנמדדו באותן השנים בתחנה מטאורולוגית סמוכה, דבר המצביע על כך שלפלסטיות פנוטיפית (תגובה לתנאי סביבה משתנים) יש חלק בהסבר של השונות הפנולוגית של העצים. נמצא גם מתאם גבוה בין ערכי המשתנים הפנולוגיים של כל עץ בין השנים כלומר למרות השינויים בין השנים וההבדלים בין האוכלוסיות, כאשר לכל עץ קיים דגם קבוע. כלומר, בכל אוכלוסייה יש עצים שהם תמיד הראשונים להשיר עליהם ועצים אחרים תמיד מאחרים. דגם קבוע הזה מלמד כי השונות בהרכב הגנטי בין הפרטים היא מרכיב חשוב בהסבר השונות הפנוטיפית בפנולוגיה של העצים, כלומר ההבדלים במועדי נשירה ולבלוב של אלון התבור בישראל. נראה שבתנאי אקלים ים-תיכוניים בהם החורף מתון מאוד, לעצים ירוקי עד יש יתרון יחסית לעצים העומדים בשלכת חורף, זאת מכיוון שתקופת הטמעה שלהם ארוכה יותר בכל שנה. לאור הנחה זו ותוצאות המחקר, ניתן לשער כי עצי אלון התבור בישראל נמצאים בשלב מעבר אבולוציוני משלכת חורף לירוקי עד. השערה זו תואמת את היות הסקציה (קבוצת המינים) לה שייך אלון התבור קבוצה של מינים נשירים המותאמים להרים גבוהים, כאשר אלון תבור הוא מין שהרחיב את תפוצתו למעונות נמוכים וחמים. שינוי זה הוא כנראה הגורם לכך שהוא, כנראה, נמצא כעת בשלב מעבר אבולוציוני ממין העומד בשלכת חורף למין ירוק-עד.
מבוא
שלכת, שהיא תרדמה של צמחים המלווה בהשרה סינכרונית של כל העלים התפתחה בצמחים כהתאמה לתנאי עקה קיצונית (בעיקר יובש במקומות חמים ובמקומות קפואים) תוך צמצום מוחלט של שטח העלים שדרכם מתבצע עיקר נידוף ואיבוד המים בצמחים (Axelrod, 1966; Kozlowski, 1976).עצים נשירי חורף הם צורת-חיים אופיינית לאזורים צפוניים בעלי אקלים ממוזג בעולם ובחגורת הביניים בהרים גבוהים, בהם החורף קפוא והעלים נמצאים בסכנת קיפאון ומוות. כמו כן, השרת עלים מצמצמת מאוד את הנזקים הנגרמים מעומס של שלג הנערם על ענפים נושאי עלים ושובר אותם. לצמחים העומדים בשלכת ונמצאים בתרדמה ללא עלים בתקופות קשות יש יתרונות וחסרונות יחסית לעצים ירוקי עד הנושאים את עליהם בכל ימות השנה. היתרונות היחסיים של כל צורת-חיים תלויים בתנאי בית הגידול ולכן הם גם משפיעים על בתי הגידול והתפוצה הגיאוגרפית העולמית שלהם (Moore, 1980 ;Chabot and Hicks, 1982; Kikuzawa,1989).
עצים ושיחים ירוקי עד בעלי עלים גלדניים המותאמים לתנאי יובש הם צורת החיים העיקריות האופייניות לחמשת האזורים בעלי אקלים ים-תיכוניים בעולם הם (Naveh, 1967; Mooney and Dunn, 1970a; Raven, 1971; Aschman, 1973; Cody and Mooney, 1978; Orshan, 1986, 1989). משך החיים של כל עלה בודד ארוך יחסית במינים אלה מאשר במינים העומדים בשלכת (Moore, 1980; Field and Mooney, 1983). טמפרטורות נמוכות, או משך יום קצר מצמצמים באופן קיצוני את ההפרש בין שיעור ההטמעה לנשימה ברמת העלה, ואם עלה הופך מיצרן לצרכן הוא נושר מהעץ (Larcher and Bauer, 1981). החורף באקלים ים-תיכוני הוא גשום, אבל הטמפרטורות אינן יורדות בדרך כלל מתחת לנקודת הקיפאון (Mooney et al., 1974; Quezel, 1985). בתנאים אלה יכולים צמחים להטמיע ולצמוח גם בחורף, כפי שעשים הצמחים העשבוניים החד-שנתיים בישראל הנובטים בסתיו, צומחים בחורף ופורחים באביב המוקדם. בצמחים המעוצים של החורש (צ'פרל) בקליפורניה, שגם בה יש אקלים ים תיכוני, נמצא ששיעור ההטמעה גבוה מהנשימה גם בחורף (Mooney and Dunn, 1970a,b). באגן הים התיכון נמצא שקצב ההטמעה במינים שונים: אלונים (Quercus ilex, Q. pubescens), זית אירופי (Olea europea) וקטלב (Arbutus unedo) בחורף נמוך רק במעט מאשר בקיץ (Larcher and Bauer, 1981,Beyschlag et al., 1990). בלוטם מרווני (Cistus salvifolius) המחליף עלי קיץ בעלי חורף, ההטמעה בחורף גבוהה יותר מאשר בקיץ (Harley et al., 1987). הקמביום היא רקמה מריסטמטית האחראית על ההתעבות המשנית והצמיחה לרוחב של גזעי עצים. בעצים נשירים מתחילה פעילות הקמביום רק לאחר החורף במקביל ללבלוב האביבי, לעומת זאת בעצים ירוקי עד ים תיכוניים יש שהיא נמשכת גם בחורף (Liphschitz and Lev-Yadun, 1986). בחבל הים-תיכוני בישראל, החורף הוא עונת הצמיחה העיקרית של העשבוניים החד-שנתיים שרבים מהם נובטים בסתיו וגדלים בחורף בזמן שעצים נשירי חורף נמצאים בתרדמה. לאור כל האמור, ניתן להניח שלעצים ירוקי עד יש יתרון יחסית לעצים נשירי חורף החולקים איתם את בית הגידול. לכן ניתן גם לצפות שהארכת משך חיי העלים וצמצום משך השלכת תקנה יתרון אדפטיבי לעצים במינים נשירי חורף באגן הים התיכון.
לעצים ירוקי הגדלים בקרקעות דלות מינרלים בהן רמות נוטריינטים נמוכות מגבילות את קצב גידול העלים, יש יתרון יחסית לעצים נשירים (Jonasson, 1989; Goldberg, 1982; Chabot and Hicks, 1982). בהתאמה לכך נמצא שעצים ירוקי עד נפוצים יותר בבתי גידול בהם העקה גדולה ועצים נשירים נפוצים יותר בבתי גידול נוחים יותר (Kikuzawa, 1989). מכיוון שהקרקעות הים תיכוניות הן דלות מינרלים, צפוי שלעצים ירוקי עד ים תיכוניים יהיה יתרון יחסי לעומת עצים נשירי חורף (Shaver, 1981).
למרות האמור, כ 45% מיני העצים הגדלים בחבל הים-תיכוני בישראל הם נשירי חורף (שמידע 1983). תופעה זו מוסברת על ידי נדידה של מיני צמחים מאזורים פיטוגאוגרפיים סמוכים בהם החורף קפוא (Danin and Plitman, 1989), או כתוצאה מההשפעה של תקופות קרחוניות (Shmida, 1978; Zohary, 1973; Pons and Quezel, 1985). בספרד נמצא שמקורם של מיני העצים ירוקי העד בעלי עלים גלדניים בתחילת עידן הפלייסטוקן בזמן שרר שם אקלים טרופי. לאחר מכן, מינים אלה פתחו מנגנונים לעמידות בפני יובש המאפשרים להם לשרוד בתנאים הנוכחיים של הקיץ הים-תיכוני (Herrera 1992).
אלון התבור הוא מין המוגדר כעומד בשלכת חורף (Zohary, 1973). מכל האמור לעיל, ניתן להניח שבבית גידולו הטבעי בעמקים ובגבעות נמוכות במזרח אגן הים התיכון לפרטים ירוקי עד צפוי להיות יתרון יחסי לעומת פרטים בעלי שלכת חורף ארוכה.
מטרת מחקר זה הייתה לבדוק האם קיימת חוקיות בפנולוגיה של השרת העלים באלון התבור ולבדוק האם יתכן שמין זה נמצא בתהליך מעבר אבולוציוני ממין בעל שלכת חורף למין ירוק עד.במידה והתוצאות תתמוכנה בהשערה זו, זה יכול להיות הסבר נוסף למקורם של חלק ממיני העצים ירוקי עד באגן הים התיכון. כלומר שיתכן שיש מינים שלא התפתחו מינים טרופיים ירוקי עד שעברו התאמה ליובש בקיץ הים תיכוני, אלא התפתחו כאן ממינים נשירי חורף שהיו כאן בתקופות הקרח, או שחדרו לאזור הים-תיכוני מאזורים פיטוגאוגרפיים סמוכים בהם החורף קפוא.
חומרים ושיטות
אלון התבור
אלון התבור(Quercus ithaburensis Decne) הוא הנציג המזרח ים-תיכוני של קבוצת אלוני aegilops (לפי Zohary, 1973). זהרי הגדיר בישראל חמישה תת-מינים של אלון התבור וכל האוכלוסיות במחקר זה שייכות לתת המין תבור (var. ithaburensis). בטורקיה גדל תת-מין אלון תבור גדול קשקשים (var. macrolepis) ונמצאו הבדלים מורפולוגיים בולטים בין שני תת-המינים (Dufour-Dror and Ertas, 2002) כמו גם בפרמטרים אקלימיים שונים המאפיינים את אזורי התפוצה של כל אחד מהם (Dufour‐Dror and Ertas, 2004).) הירקון מהווה את גבול התפוצה הדרומי העולמי של אלון התבור, שהוא מין תרמופילי (חובב חום) וגדל בדרך כלל ברום נמוך מ 500 מ' מעל לפני הים (Zohary, 1973) ורק במקרים נדירים עד 1000 מ' מעל לפני הים (Kaplan, 1984).,אלון התבור גדל על מגוון של קרקעות (קרקע סחף, חול, רנדזינה ובזלת), בגליל התחתון הוא גדל במפנים הדרומיים בעת שאלון מצוי גדל על המפנים הצפוניים של הנחלים (Aloni and Orshan, 1972; הר ,2008; הר ושות, 2015). בחודשי הקיץ נמדדו באלון התבור ערכים גבוהים של נידוף מים ושורשיו מגיעים לקרקע עמוקה הנשארת לחה גם בקיץ (Schiller et al. 2007). עצי אלון התבור הגדלים על גבעות קרטוניות מחדירים את שורשיהם לכיסי קרקע המספקים להם את כמות המים הדרושה להם בדיפוזיה מהסלע (הר, 2008).
מכל העצים הדומיננטיים המכסים שטחים נרחבים בחבל הים-תיכוני בישראל, אלון התבור הוא היחיד העומד בשלכת חורף (Zohary, 1973). יערות פתוחים של אלון התבור היו נפוצים בעבר בכל העמקים הגדולים בצפון ישראל ולאורך מישור החוף (Eig 1933; Zohary, 1973), חלקם נכרתו להכשרת שטחי חקלאות והנותרים נכרתו בתחילת המאה העשרים על ידי התורכים לספק דלק לקטרים במלחמת העולם הראשונה (פז וזחלקה, 1997).


מימין: אלוני תבור בשלכת בחלקת המחקר באלונים, צילם: עוזי פז ©; משמאל: אלוני תבור מלבלבים בחלקת המחקר באלונים, צילם: עוזי פז ©
להגדלה – לחצו על התמונות
מספר חוקרים שמו לב לשונות המורפולוגית הגבוהה בעצי אלון התבור (Zohary, 1973; Kaplan, 1984). זהרי (Zohary, 1973) הגדיר את אלון התבור כמין בעל שלכת לא סדירה וציין כי יש עצים שלעתים בחורפים מתונים אינם משירים את כל עליהם ('It may fail to shed its leaves in years of mild winters'), ברור סוגיה זו היא נושא המחקר הנוכחי.
אתרי המחקר
המחקר בוצע בארבעה אתרים: חדרה, אלונים, בית רימון ויהודיה (איור 1) שבכל אחד מהם קיימת אוכלוסייה אחידה של עצים בוגרים שלאלון התבור ותנאי סביבה אחידים. כל אתר מהאתרים מייצג אוכלוסייה אופיינית של אלון התבור בצפון ישראל במרחקים שונים מהים, ברום ותנאי סביבה מעט שונים (טבלה 1). בכל אתר נבחרו באקראי כ 60 עצים בוגרים. בנוסף לכך נבחרו עצים נוספים בגליל התחתון שאחוז כיסוי העלים שלהם בסוף החורף היה גבוה במיוחד, קבוצה זו קיבלה את השם 'ירוקי עד'.

איור 1. מפת תפוצה של אלון התבור בישראל. האותיות מסמנות את מיקום אוכלוסיות המחקר: חדרה (H), אלונים (A), בית רימון (R), ויהודייה (Y).
להגדלה – לחצו על התמונה
טבלה 1. אתרי המחקר של אלוני התבור והתחנה המטאורולוגית הסמוכה (בסוגריים), מיקומם הגיאוגרפי, סוג הקרקע, המרחק מהים, הרום, כמות המשקעים השנתית הממוצעת והטמפרטורות הממוצעות בחורף ובקיץ.
| אתר | איזור | מיקום | קרקע |
מרחק מהים (ק"מ) |
רום ( מ') |
משקעים (מ"מ לשנה) |
טמפרטורה ממוצעת (C°) |
|
| ינואר | אוגוסט | |||||||
| חדרה (זכרון יעקב) | מישור החוף |
32.45°N 34.96°E |
חול | 10 | 50 | 556 | 12.8 | 25.9 |
| אלונים (נווה יער) | גליל תחתון |
31.72°N 35.18°E |
אלוביאלית | 35 | 100 | 578 | 12.1 | 26.9 |
| בית רימון (כדורי) | גליל תחתון |
32.77°N 35.29°E |
אלוביאלית | 50 | 100 | 521 | 11.4 | 26.6 |
|
יהודיה (קצרין) |
גולן | 32.93°N 35.69°E | גרומוסול | 90 | 550 | 550 | 8.6 | 24.5 |


מימין: אלוני תבור באוכלוסיית חדרה בשלכת חלקית, צילם: עוזי פז ©; משמאל: אלון תבור 'ירוק עד', העץ מלבלב כשהוא עדיין נושא את העלים של השנה הקודמת, צילם: עוזי פז ©
להגדלה – לחצו על התמונות
תצפיות
כל אחד מהעצים סומן במספר ובוקר כל 4-2 שבועות החל מתחילת אוקטובר (מועד תחילת השרת העלים) בסתיו ועד השלמת הלבלוב בסוף מרס. התצפיות החלו בסתיו 1988 ונמשכו עד אביב 1992. באוכלוסיית חדרה בוצע הניטור במשך שתי העונות הראשונות בלבד. בכל ביקור בוצעה הערכה חזותית של אחוז העלים הבוגרים ואחוז העלים הצעירים על כל עץ. העלים הבוגרים היו אלה שהופיעו על העץ באביב הקודם או בקיץ, העלים הצעירים היו אלה שהופיעו באביב לאחר שהעץ עמד בשלכת. דרגות כיסוי העלים היו: 95% (תחילת נשירת עלים חומים), 75%, 50%, 25%, ו 5% (רק עלים בודדים על העץ).
מועד התצפיות מציין את מספר השבוע כאשר השבוע הראשון של אוקטובר בכל שנה היה שבוע מספר אחד. המשתנים הפנולוגיים שנקבעו היו: מועד תחילת השלכת: מספר השבוע בו נצפתה תחילת השרת עלים. משך השרת העלים: מספר השבועות שחלף ממועד תחילת השלכת עד השבוע בו נצפו רק מעט עלים בוגרים על העץ. מועד תחילת הלבלוב : מספר השבוע בו נצפו ניצנים פתוחים והעלים הראשונים הופיעו. משך השלכת: מספר השבועות בו עמד העץ ללא עלים, ממועד סיום השרת העלים עד מועד פתיחת הניצנים ותחילת הלבלוב. משך הלבלוב: מספר השבועות ממועד תחילת הלבלוב ועד המועד בו כל העץ כוסה בעלים צעירים.
ניתוח התוצאות
תחילה נקבעו כל המשתנים הפנולוגיים לכל עץ. במידה ושלב מסוים לא נצפה במדויק הוא נקבע על ידי אקסטרפולציה של גרף אחוז העלים כנגד מספר השבוע. ניתוח שונויות דו-כיווני (Two-way ANOVA) בוצע להערכת ההשפעה של האתר ושנת התצפית על המשתנים הפנולוגיים ובמיוחד על משך השלכת. בכל מקרה בו מתגלה אינטראקציה בין הגורמים המשפיעים לא ניתן לבדוק את ההשפעה של כל גורם בנפרד (Underwood, 1981). לכן לא מוצגת ההשפעה של כל גורם בנפרד (one-way ANOVA) ולא השוואה של הערכים הממוצעים בין האוכלוסיות והממוצעים בין השנים.
נמצא מתאם גבוה ומובהק בין מרבית המשתנים הפנולוגיים, לכן לא כל המשתנים מוצגים בכל טבלאות והאיורים. רגרסיה מרובה בשלבים בוצעה לבדיקת הגורמים העיקריים שהשפיעו על משך השלכת. מקדם המתאם (קורלציה) של Pearson חושב לגבי המשתנים הפנולוגיים של כל עץ בשנים השונות, כמו כן נבדקה הקורלציה בין המשתנים הפנולוגיים לבין נתוני מזג האוויר בשנים הרלוונטיות שהתקבלו ומהשרות המטאורולוגי לגבי התחנות הקרובות ביותר לאתרי המחקר.
האנליזות הסטטיסטיות בוצעו בעזרת הפרוצדורות: GLM ,CORR ,REG בתכנת SAS (SAS Institute 1988).
תוצאות
פנולוגיה של העלווה ברמת העץ הבודד
נמצאו הבדלים גדולים בפנולוגיה של העלווה בין פרטים שונים. בפרטים מסוימים השרת העלים התחילה כבר בשבוע הראשון של אוקטובר ואילו בפרטים אחרים השרת העלים התחילה רק בתחילת מרס. להדגמת ההבדלים הגדולים, באיור 2 מוצגת ההתנהגות העונתית של העלווה בשני עצים קיצוניים. בעץ הראשון (יהודייה 108 בחורף 1991-2) שהשרת העלים הסתיימה 14 שבועות לפני תחילת הלבלוב, כלומר משך השלכת היה 14 שבועות, ולגרף המתאר את השינויים באחוז כיסוי העלווה בשמך העונה צורה של U (איור 2). לעומת זאת בעץ השני (חדרה 515 בחורף 1989-90) השרת העלים הסתיימה11 שבועות לאחר תחילת הלבלוב האביבי ולגרף צורה של X. מהגרף עולה כי אחוז הכיסוי של העלווה היה תמיד גבוה מ 10% ולכן ניתן להגדירו כירוק עד (איור 2). עצים דומים בהם סיום השלכת היה לאחר תחילת הלבלוב האביבי נמצאו בכל אתרי המחקר והיוו 1.7% בבית רימון, 14.6% ביהודייה, 14.7% באלונים ו 41% בחדרה ו-18% בממוצע מכלל העצים שנבדקו. לאחוז זה היה מתאם שלילי עם משך השלכת הממוצע בכל אוכלוסייה. עבור 39 מתוך 42 משתנים, נמצאה קורלציה מובהקת בין זוגות שנים, כולל משך השלכת שהוא המשתנה החשוב ביותר (טבלה 2).

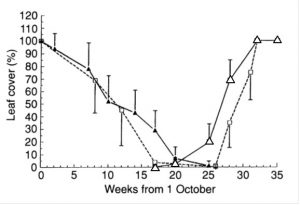
איור 2 (מימין) – השינויים באחוז הכיסוי של עלים בוגרים וצעירים בעץ 'יהודייה 460' בחורף 1991-2, של עלים בוגרים וצעירים בעץ 'חדרה 515' בחורף 1989-90. הציר האופקי מציין את מספר השבועות שחלפו מהראשון באוקטובר בכל שנה.
איור 3 (משמאל) – השינויים באחוז הכיסוי הממוצע (± סטיית התקן) של עלים בוגרים וצעירים באוכלוסיית בית רימון בחורף 1991-2, ושל עלים בוגרים וצעירים באוכלוסיית חדרה וחדרה בחורף 1989-90. הציר האופקי מציין את מספר השבועות שחלפו מהראשון באוקטובר בכל שנה.
לצפייה מיטבית – לחצו על התמונות
טבלה 2. מקדם המתאם פירסון (r) בין המשתנים הפנולוגיים: תחילת השרת העלים (תהע), משך השרת העלים (מהש), סוף השרת העלים (סהע), משך השלכת (מש), תחילת הלבלוב האביבי (תל), משך הלבלוב האביבי (מל) וסוף הלבלוב האביב (סל) של עצי אלון התבור בכל אתרי המחקר בין כל זוגות השנים 89 (n=248) ו-90 (n=248), והשנים 91 (n=188) ו-92 (n=190). האות n מסמנת את מספר העצים בכל שנה ומספר הכוכביות מציין את דרגת המובהקות הסטטיסטית: לא מובהק=ns, *P<0.05, **P<0.01,***P<0.001 .
| תהע 89 | תהע 90 | תהע 91 | מהע 89 | מהע 90 | מהע 91 | ||
| תהע 90 | 0.74*** | מהע 90 | 0.48*** | ||||
| תהע 91 | 0.57*** | 0.62*** | מהע 91 | 0.24** | 0.34** | ||
| תהע 92 | 0.59*** | 0.60*** | 0.78*** | מהע 92 | 0.12ns | 0.24* | 0.34*** |
| סהע 89 | סהע 90 | סהע 91 | מש 89 | מש 90 | מש 91 | ||
| סהע 09 | 0.43*** | מש 90 | 0.64*** | ||||
| סהע 91 | 0.34*** | 0.34*** | מש91 | 0.22** | 0.26** | ||
| סהע 92 | 0.35*** | 0.38*** | 0.51*** | מש 92 | 0.36*** | 0.42*** | 0.55*** |
| תל 89 | תל 90 | תל 91 | מל 89 | מל 90 | מל 91 | ||
| תל 90 | 0.47*** | מל 90 | 0.28*** | ||||
| תל 91 | 0.64*** | 0.74*** | מל 91 | -0.09 | -0.11ns | ||
| תל 92 | 0.22** | 0.38*** | 0.41*** | מל 92 | 0.20** | 0.31*** | 0.11ns |
| סל 89 | סל 90 | סל 91 | |||||
| סל 90 | 0.525*** | ||||||
| סל 91 | 0.63*** | 0.63*** | |||||
| סל 92 | 0.40*** | 0.48*** | 0.72*** |
פנולוגיה של העלווה ברמת האוכלוסייה
הגרף המתאר את ההתנהגות העונתית של העלווה ברמת האוכלוסייה (איור 3) דומה לזו ברמת העץ הבודד (איור 2). בבית רימון בחורף 1991-2 השרת העלים הסתיימה לפני שהחל הלבלוב האביבי ולכן לעקומה צורת U, לעומת זאת בחדרה בחורף 1989-90 השרת העלים הסתיימה רק לאחר שהתחיל הלבלוב האביבי וכתוצאה מכך לעקומה צורת X. העקומות של האוכלוסיות באלונים ויהודייה היו בעל דגם ביניים (איור 3). נמצאו הבדלים גדולים בין כל המשתנים הפנולוגיים בין האוכלוסיות ובין השנים (איור 4). בניתוח שונויות דו-כיווני נמצאה השפעה מובהקת של האתר, של השנה וגם האינטראקציה ביניהם הייתה מובהקת (טבלה 3). פרוש הדבר שההשפעה של השנה הייתה שונה באוכלוסיות שונות ולכן לא ניתן לקבוע האם האתר או השנה הם הפקטור העיקרי המשפיע על הפנולוגיה של העלווה באלון התבור.

איור 4. הערכים הממוצעים (וסטיית התקן) של מועד תחילת השרת העלים (1) משך השרת העלים (2), משך השלכת (3) ומשך הלבלוב האביבי (4) בעצי אלון התבור באוכלוסיות המחקר: יהודייה (Y), בית רימון (B), אלונים (A), חדרה (H) ובקבוצת ירוקי העד (E) בחורף בשנים 1988-89 , 1989-90 , 1990-91 , 1991-2 . המועדים הם מספרי השבועות לאחר הראשון באוקטובר בכל שנה.
טבלה 3. סיכום התוצאות (Mean squares), דרגות החופש (df)) והפרופורציה של השונות המוסברת (R2) של מבחני ניתוח שונויות דו-כיווני (Tow way ANOVA) לבדיקת ההשפעה של האוכלוסייה (חדרה, אלונים, בית רימון, יהודייה וירוקי עד) , השנה והאינטראקציה ביניהם על המשתנים הפנולוגיים הממוצעים של עצי אלון התבור בכל אתרי המחקר ובמשך כל שנות המחקר. כל התוצאות מובהקות מאוד (P<0.01).
| מקור השונות | Df | תחילת השרת עלים | משך השרת העלים | סוף השרת העלים | משך השלכת | תחילת הלבלוב | משך הלבלוב |
| R2 | 0.62 | 0.42 | 0.50 | 0.57 | 0.76 | 0.66 | |
| Model | 17 | 457 | 283 | 300 | 507 | 365 | 250 |
| אוכלוסיה | 4 | 1498 | 662 | 469 | 1075 | 933 | 355 |
| שנה | 3 | 414 | 551 | 856 | 1188 | 245 | 778 |
| שנה X אוכלוסייה | 10 | 53 | 51 | 65 | 76 | 173 | 49 |
בבית רימון השלכת התחילה מאוחר יחסית לשאר האתרים, אולם קצב השרת העלים היה מהיר ולכן משך השלכת היה הארוך ביותר ובחדרה היה משך השלכת הקצר ביותר. ההבדלים במועד תחילת הלבלוב האביבי ומשכו היו קטנים יותר מאשר ההבדלים במועדי תחילת השרת העלים ומשך ההשרה (איור 4). מקדם השונות (סטיית התקן חלקי הממוצע) של משך השלכת בין השנים היה נמוך יותר בבית רימון (37.8%) מאשר בשאר האתרים (74.1-207.3%). מקדם השונות של הממוצע הכללי של כל האוכלוסיות במשך כל שנות התצפית של מועד תחילת הלבלוב (13.2%) וסיומו (6.7%) היו קטנים יותר מאשר מועד תחילת השרת העלים (50.8%) וסיומה (16.8%).
הקורלציה בין כל זוגות המשתנים הפנולוגיים של כל העצים במשך כל השנים (n=874) הייתה מובהקת פרט לזו שבין מועד סיום השרת העלים ומועד סיום הלבלוב. משך השלכת היה בקורלציה הגבוהה ביותר עם מועד סיום השרת העלים ומועד הלבלוב (טבלה 4).
טבלה 4. מקדם המתאם של פירסון (r) בין זוגות של המשתנים הפנולוגיים של כל עץ בכל האוכלוסיות בכל השנים (n=874) . כל הקורלציות מובהקות: *p<0.05, **P<0.01. ***P<0.001.
| סוף לבלוב | משך לבלוב | תחילת לבלוב | משך שלכת | סוף השרת עלים | משך השרת עלים | |
| תחילת השרת עלים | 0.28*** | -0.28*** | 0.43*** | -0.08* | 0.49*** | -0.58*** |
| משך השרת עלים | -0.243*** | 0.16*** | -0.28*** | -0.54*** | 0.41*** | |
| סוף השרת עלים | 0.06 | -0.15*** | 0.17*** | -0.69*** | ||
| משך שלכת | 0.32*** | -0.46*** | 0.60*** | |||
| תחילת לבלוב | 0.49*** | -0.80*** | ||||
| משך לבלוב | 0.13*** |
הקורלציה בין ממוצעים של מועדי תחילת השלכת וסיומה ושל משך השלכת של האוכלוסיות בכל שנה וטמפרטורת המינימום היומי הממוצע והטמפרטורות המקסימליות בין החודשים ספטמבר ואפריל מוצגים בטבלה 5. נמצאה קורלציה שלילית מובהקת בין מועד סיום השרת העלים ומשך השלכת לבין הטמפרטורות היומית המרבית הממוצעות בחודשים דצמבר ינואר ופברואר. מועד תחילת השרת העלים היה בקורלציה עם מספר נתונים אקלימיים של חודש הסתיו (טבלה 5). לעומת זאת לא נמצאה קורלציה בין המשתנים הפנולוגיים לבין כמות המשקעים בשנה החולפת.
טבלה 5. מקדם המתאם של פירסון (r) בין מועד תחילת השרת העלים, סיומו ומשך השלכת הממוצע בכל האוכלוסיות לבין טמפרטורות המינימום (מינ.) והמקסימום (מקס.) בחודשים ספטמבר עד אפריל, שנמדדו בתחנות מטאורולוגיות סמוכות. קורלציות מובהקות מסומנות על-ידי כוכביות: לא מובהק=ns, *p<0.05, **P<0.01 ***P<0.001.
| משתנה | משך השלכת | סוף השרת עלים | תחילת השרת עלים |
| ספטמבר מינ. | 0.13 | 0.32ns | 0.6* |
| ספטמבר מקס. | -0.11 ns | -0.03 ns | 0.16 ns |
| אוקטובר מינ. | 0.32 ns | 0.18 ns | 0.59* |
| אוקטובר מקס. | 0.39 ns | 0.14 ns | 0.58* |
| נובמבר מינ. | -0.08 ns | 0.58 ns | 0.35 ns |
| נובמבר מקס. | -0.21ns | 0.71** | 0.57* |
| דצמבר מינ. | -0.16 ns | 0.43 ns | 0.43 ns |
| דצמבר מקס. | -0.64* | 0.79*** | 0.55* |
| ינואר מינ. | -0.54 ns | 0.74** | 0.40 ns |
| ינואר מקס. | -0.53* | 0.76** | 0.52 ns |
| פברואר מינ. | -0.17 ns | 0.75** | 0.55* |
| פברואר מקס. | -0.70** | 0.65* | 0.44 ns |
| מרס מינ. | -0.23 ns | 0.71** | 0.73** |
| מרס מקס. | 0.38 ns | 0,69** | 0.38 ns |
| אפריל מינ. | 0.82** | 0.53* | -0.01 ns |
| אפריל מקס. | 0.20 ns | 0.07 ns | -0.33 ns |
דיון
במחקר זה נמצאו הבדלים ניכרים בדגם ההתנהגות העונתית של העלווה, בעיקר במועדי תחילת השרת העלים בסתיו, בין האתרים ובין שנות התצפית (איור 4). הבדלים אלה יכולים להיות תוצאה של הבדלים בהרכב הגנטי של האוכלוסיות או השפעה של ההבדלים בתנאי הסביבה בה גדלות האוכלוסיות האלה. אולם משום שהייתה אינטראקציה סטטיסטית מובהקת בין השפעת האתרים לבין השפעת השנים (טבלה 3), כלומר תנאי הסביבה השפיעו בצורה שונה על האוכלוסיות השונות, לא ניתן לקבוע האם האתר או השנה הם הפקטור העיקרי המשפיע על הפנולוגיה של העלווה באלון התבור.
בחדרה, גדלה האוכלוסייה ברום הנמוך ביותר והקרבה המרבית לים, תנאי האקלים הם המתונים ביותר, הקיץ פחות חם והחורף פחות קר (טבלה 1). משך השלכת הממוצע בחדרה היה הקצר ביותר ואחוז העצים 'ירוקי העד' שסיימו את השרת העלים שלהם רק לאחר תחילת הלבלוב האביב היה הגדול ביותר יחסית לכל שאר אוכלוסיות המחקר. לעומת זאת, באוכלוסיית בית רימון היה משך השלכת הארוך ביותר ואחוז העצים 'ירוקי העד' בה היה הנמוך ביותר מבין כל האוכלוסיות הנחקרות. באוכלוסיות אלונים ויהודייה היה דגם ביניים במרבית המשתנים שנבדקו, זאת למרות שאתר אלונים קרוב לבית רימון וביהודייה ערכי האקלים בחורף קיצוניים יותר. הדגם הקיצוני יותר בבית רימון נובע כנראה מהתנאים המקומיים, שכן האתר נמצא בעמק בין הרים המנקז לתוכו אויר קר בחורף ולעיתים תכופות יש שם קרה בלילה. אולם עובדה זו אינה משתקפת בנתונים האקלימיים שהתקבלו מתחנה המטאורולוגית הקרובה (טבלה 1).
כאמור, ההבדלים הפנולוגיים בין האוכלוסיות הגדלות באתרים שונים יכולים להיות תוצאה של הבדלים גנטיים בין האוכלוסיות ויכולים להיות תוצאה של פלסטיות פנוטיפית, כלומר לשקף את ההשפעה של תנאי הסביבה כגון סוג קרקע ותנאי אקלים, זאת גם בהעדר הבדלים גנטיים בין האוכלוסיות. למרות שפלסטיות פנוטיפית משקפת את השפעת תנאי הסביבה, טווח הפלסטיות הפנוטיפית שונה במינים ביולוגיים שונים, היא עצמה תכונה נורשת והיא מגדילה את השונות של הפנוטיפים הנתונים לסלקציה מעבר למספר הגנוטיפים הקיים (West-Eberhard, 1989). סיווג מקור השונות כגנוטיפי או פלסטיות פנוטיפית יכול להיות מבוצע באופן ניסויי בו שותלים גנוטיפ מסוים במקומות עם תנאי סביבה שונים. אולם זה כמעט שלא ניתן לביצוע עם עצים מאריכי ימים ויש לנסות ולהשתמש בשיטות אחרות.
שונות גנטית נמצאה בין אוכלוסיות של אלון אילקס (Q. ilex) בצרפת (Yacine and Lumart, 1988), ואוכלוסיות של אלון רובור (Q. robur), אך למרות זאת, שלא כמו באלון התבור, במשך שבע שנות מחקר לא נמצאו הבדלים פנולוגיים בין פרטים ובין אוכלוסיות (Crawley and Akhteruzzaman, 1988).
בסיכום מחקרו על אוכלוסיית אלון התבור ביער יהודייה כותב דידי קפלן (Kaplan, 1984): "עצים יכולים להימצא בשלבי מעבר שונים, תופעה שהיא אחד הביטויים של השונות הגנטית באלון התבור". הבדלים בגודל ובצורה של הספלול והקשקשים תוארו באלון התבור והיו הבסיס לחלוקתו לתת-מינים (Zohary, 1972). שונות כזו יכולה לשקף שונות גנטית בין פרטים ובין אוכלוסיות, אבל יכולה להיות גם תוצאה של פלסטיות פנוטיפית.
אוכלוסיות אלון התבור שנבחרו למחקר זה היו הומוגניות מבחינת הרום, הקרקע, המפנה והשיפוע על מנת לצמצם את ההבדלים הסביבתיים בין הפרטים בכל אוכלוסייה, כולל בבית השורשים שבקרקע. בהעדר הבדלים סביבתיים, שונות גנוטיפית יכולה להסביר את ההבדלים הגדולים שנצפו בין פרטים באותה אוכלוסייה. לעיתים קרובות ניתן לראות בחורף שני עצים שכנים הגדלים על קרקע סחף אחידה ועמוקה כאשר אחד עומד בשלכת מלאה ושכנו עדיין מכוסה בעלווה, והבדל זה חוזר על עצמו מידי שנה ללא תלות בטמפרטורות וכמות המשקעים.
השוואה של המשתנים הפנולוגיים של עצים בודדים לאורך השנים מגלה קורלציה גבוהה בעיקר מועד תחילת השרת העלים ומועד סיומה, זאת למרות השינויים ברמת האוכלוסייה בין השנים ובין הפרטים בתוך האוכלוסייה בכל שנה נתונה (טבלה 2). כלומר, פרטים שלהם משך שלכת ארוכה יחסית לפרטים אחרים ישמרו על מיקומם היחסי בכל השנים, כאלה שבהם החורף מתון או קר יותר. עובדה זו מחזקת את האפשרות שההבדלים הפנולוגיים בין הפרטים מתבססים על הבדל בין הגנוטיפים של הפרטים. לעומת זאת השינויים במשתנים הפנולוגיים של כל פרט ושל האוכלוסייה משנה לשנה והקורלציה שלהם עם הטמפרטורות בסתיו ובחורף (טבלה 5) מרמזים כנראה על השפעה סביבתית באמצעות פלסטיות פנוטיפית.
משך החיים של עלה בודד בעצים העומדים בשלכת הוא מן הסתם קצר מ 12 חודשים (Kikuzawa, 1989, 1991). מועד הלבלוב האביבי ומועד השרת העלים בסתיו מווסת על-ידי גורמים סביבתיים בעיקר משך היום, הטמפרטורות ומגמת השינויים בהם כלומר התארכות היום או התקצרותו, התחממות או התקררות (,Salisbury 1981). סלקציה עשויה לשנות את משך תקופת השלכת על-ידי דחייה של השרת העלים בסתיו, או על-ידי הקדמת הלבלוב האביבי, אם זה מקנה יתרון לפרט ואם תנאי הסביבה מאפשרים זאת. תהליך כזה יכול להסתיים בדחיה של השרת העלים עד לאחר הלבלוב האביבי ואם זה קורה, העץ ייחשב כירוק עד משום שהוא נושא עלים במשך כל השנה.
על פי החלוקה של (Kikuzawa, 1983, 1984) הלבלוב באלון התבור מסווג כבינוני מכיוון שבמשך כחודש העץ מתכסה בעלים באביב (Kaplan, 1984). טיפוס כזה של לבלוב נחשב כמייטבי לעצים בעלי צמיחה קבועה בסביבה יציבה (Iwasa and Cohen, 1989) . גל נוסף של צמיחה מאפיין את אלון התבור באוגוסט וספטמבר. הוא מלווה בהתארכות גבעולים ויצירה של עלים חדשים, גל זה חזק במיוחד כתגובה להרביווריה על-ידי טוואי האלון (Eriogasterph ilippsi) או לאחר שריפה (Kaplan, 1984). סוג לבלוב כזה נחשב כיתרון לעצים הגדלים בסביבה לא יציבה (Iwasa and Cohen, 1989). מועד תחילת הלבלוב האביבי ומשכו הן תכונות יציבות יחסית באלון התבור והשונות בהן נמוכה יחסית לתהליך השרת העלים בסתיו.
לעומת זאת, נמצאה שונות גדולה מאוד במועד תחילת השרת העלים ומשך השרת העלים בין אוכלוסיות ובין פרטים של אלון התבור. בניגוד לאלון רובור האירופי (Q. robur) בו נמצאה קורלציה בין מועד השרת העלים למועד הלבלוב (Crawley and Akhteruzzaman, 1988), לא נמצא קשר כזה באלון התבור (טבלה 5). ההבדלים במועדי השרת העלים היו גדולים בהרבה מההבדלים במועדי הלבלוב. לכן נראה שהשרת העלים באלון התבור מושפעת מתנאי הסביבה (בעיקר מהטמפרטורות בחורף) יותר מאשר הלבלוב האביבי, ובכך הוא גם חשוף יותר לברירה הטבעית (West-Eberhard, 1989). נראה שבתנאי החורף הים-תיכוני, דחייה של מועד השרת העלים מקנה יתרון לעץ בכך שהיא מעריכה את משך ההטמעה שלו. דחייה קיצונית של השרת העלים, עד לאחר מועד תחילת הלבלוב באביב, עשויה להאריך את משך החיים של מרבית העלים ל 13-12 חודשים ולהפוך את העץ מנשיר חורף לירוק עד. תוצאות המחקר תומכות בהשערה שזה הוא כנראה התהליך האבולוציוני העובר היום על אלון התבור, תהליך שבשל משך החיים הארוך של העצים עשוי להמשך מאות שנים.
הסוג אלון מונה כ 500-450 מינים הנפוצים כיום בעיקר בחציו הצפוני של כדור הארץ. מאובנים של הסוג אלון נמצאו החל מאמצע תקופת האיאוקן ומיני אלונים היו נפוצים בכל חלקי היבשת הצפונית וגם בגרינלנד(אבישי 2016). בתקופה זו שררו שם תנאים חמים ואז היו האלונים כנראה ירוקי עד ועם תהליכי התרוממות ההרים וההתקררות בצפון כדור הארץ, הם הפכו לנשירי חורף. המינים בסקצייה קבוצת המינים) בסוג אלון לה שייך אלון התבור נפוצים בעיקר בהרים גבוהים (אבישי 2016) שבהם החורף קר וקפוא ולכן הם עומדים שם בשלכת חורף. אלון תבור הוא מין מתוך קבוצה זו שהרחיב את תפוצתו למעונות נמוכים וחמים יותר, בהם שלכת חורף היא חסרון ולא יתרון. שינוי זה בתחום התפוצה הגיאוגרפית הוא כנראה הגורם לכך שאלון התבור, נמצא כנראה כעת בשלב מעבר אבולוציוני ממין העומד בשלכת חורף למין ירוק-עד.
משך החיים של העלים באלון מצוי (Q. calliprinos) שהוא ירוק עד הוא 14-13 חודשים (Orshan, 1989), הוא פורח ומלבלב בתחילת מרס ומשיר את כל העלים הישנים כחודש לאחר מכן. תופעה דומה קיימת גם בקטלב מצוי (Arbutus andrachne) ובאדר סורי (Acer obtusifoliumsyn. A. syriacum). הסוגים אדר ואלון הם סוגים צפוניים שמרבית המינים בהם נשירי חורף וכוללים גם מעט מינים ים תיכוניים שרק הם ירוקי עד. תופעה זו עשויה לרמז על כך שהמינים ירוקי העד בסוגים אלה מקורם במינים ששרדו את תקופות הקרח או שהם הגיעו לאזור הים התיכון מאזורים פיטוגאוגרפיים סמוכים (אירנו-טורני) בהם החורף קר וקפוא. לאחר מכן, בשל התנאים הנוחים בחורף הים-תיכוני ופעולת הברירה הטבעית שהקנתה יתרון לפרטים בהם השלכת קצרה יותר, הפכו להיות ירוקי עד. היפותזה זו, נוספת ומשלימה את זו הקיימת לגבי המוצא הפלאוטרופי של המינים הים-תיכוניים ירוקי העד, הטוענת שמינים אלה היו ירוקי עד בזמן ששררו תנאים טרופיים באגן הים התיכון ועם ההתייבשות הפכו עליהם להיות גלדניים והעצים עמידים ליובש (Herrera, 1992).
לסיכום, תוצאות המחקר תומכות בהשערה שאלון התבור נמצא כנראה בתהליך מעבר אבולוציוני ממין בעל שלכת חורף למין ירוק עד. השערה זו מבוססת על ההנחה שמשך חיי העלה הוא תכונה אדפטיבית (Kikuzawa, 1989, 1991; Chabot and Hicks, 1982) ושבתנאי החורף הים-תיכוני למינים ירוקי עד יש יתרון יחסי על מינים בעלי שלכת חורף (Mooney and Dunn, 1970a; Moore, 1980; Goldberg, 1982). היפותזה זו מבוססת גם על הממצאים המראים שלשונות הפנולוגית באלון התבור יש כנראה בסיס גנוטיפי ומרכיב של פלסטיות פנוטיפית ששניהם נתונים לברירה טבעית וחשובים להתחלה חיזוק וקיבוע של שינויים באוכלוסיות צמחים (West-Eberhard, 1989).
בעתיד יש עדיין לבדוק את נכונות ההשערה על-ידי מחקרים מולקולריים שיחשפו את המנגנון הגנטי, ומחקרים אקולוגיים שיראו מהו היתרון של עצים ירוקי העד לעומת עצים העומדים בשלכת האם הם צומחים יותר מהר? האם מייצרים יותר בלוטים? או שניהם יחדיו.
ספרות:
אבישי מ 2016 אלון חרמוני – מין ייחודי לחרמון. כלנית 3. https://www.kalanit.org.il/quercus-look/
פז ע וזחלקה מ 1997 היה היה יער: על איזור רמת מנשה. טבע הדברים 20: 107-87.
הר נ 2008 מבנה מערכת הסלע והקרקע והדינמיקה של משק המים בבית הגידול כגורמים אקולוגיים עיקריים בתפוצת אלון התבור והאלון המצוי באזור אלונים-מנשה. חיבור לקבלת התואר דוקטור לפילוסופיה, האוניברסיטה העברית ירושלים.
הר נ ריוב י ושני א 2015 מערך הצומח באזור אלונים-מנשה בהתאמה למבנה המרחבי של מערכת הסלע–קרקע ותכונות בתי הגידול . אקולוגיה וסביבה 6: 52-40.
שמידע א 1983 הצומח הים-תיכוני בקליפורניה ובישראל – דמיון ושוני. רתם 8: 29-5.
——————————————————————————————————–
Aloni R and Orshan G 1972 A vegetation map of the Lower-Galilee. Israel Journal of Botany 21: 209-227.
Aschman H 1973 Distribution and peculiarity of Mediterranean ecosystems. Mediterranean Type Ecosystems (eds F. di Castri and H.A. Mooney), pp. 11-19. Springer Verlag, Berlin.
Axelrod DI 1966 Origin of deciduous and evergreen habits in temperate forests. Evolution 20: 1-15.
Beyschlag B Lange OL and Tenhunen JD 1990 Photosynthesis and water relations of the Mediterranean evergreen sclerophyll Arbutus unedo L. throughout the year at a site in Portugal. III single factor analysis of light temperature and CO2 dependency of net photosynthesis. Flora 184: 271-289.
Chabot BF and Hicks DJ 1982 The ecology of leaf life spans .Annual Review of Ecology and Systematics 13: 229-259.
Cody ML and Mooney HA 1978 Convergence and dissimilarities of Mediterranean ecosystems. Annual Review of Ecology and Systematics 9: 265-321.
Crawley MJ and Akhteruzzaman M 1988 Individual variation in the phenology of oak trees and its consequences for herbivore insects. Functional Ecology 2: 409-415.
Danin A and Plitman U 1989 Revision of plant geographical territories of Israel and Sinai. Plant Systematics and Evolution 156: 43-53.
Eig A 1933 A historical phytosociological essay on Palestinian forests of Quercus aegilopsL. ssp. ithaburensis (Decne.) in past and present. Beih. Bot. Zentralbl. Abt. B. 51: 225-272. (Cited in Weisel, Pollakand Cohen 1982.)
Field C and Mooney HA 1983 Leaf age and seasonal effects on light, water and nitrogen use efficiency in a California shrub. Oecologia 56: 348-355.
Goldberg DE 1982 The distribution of evergreen and deciduous trees relative to soil type: an example from the Sierra Madre, Mexico and a general model. Ecology 63: 942-951.
Harley PC Tenhunen JD Beyschlag W and Lange OL 1987 Seasonal changes in net photosynthesis rates and photosynthetic capacity in leaves of Cistus salvifolius, a European Mediterranean semi-deciduous shrub. Oecologia 74: 380-388.
Herrera CM 1992 Historical effects and sorting processes as explanations for contemporary ecological patterns: character syndromes in Mediterranean woody plants. American Naturalist 140: 421-446.
Iwasa Y and Cohen D 1989 Optimal growth schedule of a perennial plant. American Naturalist 133: 480-505.
Jonasson S 1989 Implications of leaf longevity, leaf nutrient re-absorption and translocation for resource economy of five evergreen plant species. Oikos 56: 151-131.
Kaplan Y 1984 The ecosystem of the Yahudia Nature Reserve with emphasis on dynamics of germination and development of Quercus ithaburensis Decne. PhD thesis, University of Neimechen, the Netherlands.
Kikuzawa K 1983 Leaf survival of woody plants in deciduous broad-leaved forests. 1. Tall trees. Canadian Journal of Botany 61: 2133-2139.
Kikuzawa K 1984 Leaf survival of woody plants in deciduous broad-leaved forests. 2. Small trees and shrubs. Canadian Journal of Botany 62: 2551-2556.
Kikuzawa K 1989 Ecology and evolution of phenological pattern, leaf longevity and leaf habit. Evolutionary Trends in Plants 4: 105-110.
Kikuzawa K 1991 A cost benefit analysis of leaf habit and leaf longevity of trees and their geographical pattern. American Naturalist 138:1250-1263.
Kozlowski TT 1976 Water supply and leaf shedding. Water Deficits and Plant Growth(ed. T.T. Kozlowski), Academic Press, New York. pp. 191-231. .
Larcher W and Bauer H 1981 Ecological significance of resistance to low temperatures. Physiological Plant Ecology, Vol. 1, Responses to Physical Environment (eds. Lange O L , Nobel P S Osmond C B O. and Zelinger H) Springer Verlag, Berlin. pp. 408-439.
Liphschitz N and Lev-Ydun S 1986 Cambial activity of evergreen and seasonal dimorphics around the Mediterranean. IAWA Bulletin 7: 145-153.
Mooney HA and Dunn EL 1970a Convergent evolution of Mediterranean-climate evergreen sclerophyll shrubs. Evolution 24: 292-303.
Mooney HA and Dunn EL 1970b Photosynthetic systems of Mediterranean-climate shrubs and trees in California and Chile. American Naturalist 104: 447-453.
Mooney HA Parsons DJ and Kummerow J 1974 Plant development in Mediterranean climates. Phenology and Seasonality Modeling (ed. Leith H) Springer-Verlag, New York. pp. 265-277.
Moore PD 1980 The advantages of being evergreen. Nature 285: 535.
Naveh Z 1967 Mediterranean ecosystems and vegetation types in California and Israel. Ecology 48: 445-459.
Ne’eman G 1993 Variation in leaf phenology and habit in Quercus ithaburensis, a Mediterranean deciduous tree. Journal of Ecology 81: 627-634.
Oppenheimer HR 1947 Studies on the water balance of unirrigated woody plants. Palestine Journal of Botany Rehovot 6: 63-77.
Orshan G 1986 Plant form as describing vegetation and expressing adaptation to environment. Annali di Botanica (Rome) 44: 7-38.
Orshan G 1989 Plant Pheno-Morphological studies in the Mediterranean Type Ecosystems In: Orshan, G. (ed.) Kluwer Academic Publishers. Dordrecht, Netherlands, .2.2.
Pons A and Quezel P 1985 The history of flora and vegetation and past and present human disturbances in Mediterranean region. Plant Conservation in the Mediterranean Area (ed. Gomez-Campo C), Dr Junk Publishers, Dordrecht. pp. 25-43.
Quezel P 1985 Definition of the Mediterranean region and the origin of its flora. Plant Conservation in the Mediterranean Area (ed. C. Gomez-Campo), Dr Junk Publishers, Dordrecht. pp. 9-24.
Raven PH 1971 The relationship between Mediterranean floras. Plant Life of South-West Asia (Davis P H Harper P C and Hedge T C (eds.) Botanical Society of Edinburgh. pp. 119-134.
Salisbury FB 1981 Responses to photoperiods. Physiological Plant Ecology, Vol. 1. Responses to Physical Environment. Lange O L Nobel P S Osmond C B O and Zelinger H (eds.) SpringerVerlag, Berlin. pp. 135-139.
SAS Institute 1988 SAS/STAT user guide.
Cary NC and Shaver G 1981 Mineral nutrition and leaf longevity in anevergreen shrub Ledumpalustressp.decumbens. Oecologia 49: 362-365.
Schiller G Cohen S Ungar ED Moshe Y et al. 2007 Estimating water use of sclerophyllous species under East-Mediterranean climate: III. Tabor oak forest sap flow distribution and transpiration. Forest ecology and management 238: 147-155.
Shmida A 1978 Remarks on the paleo-climate of Sinai: based on the distribution patterns of relict plants. Prehistoric investigations in Gabel Maghara Northern Sinai. Bar-Yosef O and Phillips L (eds.) Kedem, Jerusalem. pp. 36-52.
Underwood AJ 1981 Experimental marine biology. Oceanography and Marine Biology Annual Review, 19: 513-605.
West-Eberhard MJ 1989 Phenotypic plasticity and the origins of diversity. Annual Review of Ecology and Systematics 20: 249-278.
Yacine A and Lumart R 1988 Spatial distribution of genotypes in a population of holmoak (Q. ilex L.), gene flow and mating system. Genetique Selection Evolution (France) 20: 181-198.
Zohary M 1972 Flora Palaestina, Vol. II. The Israel Academy of Sciences and Humanities, Jerusalem.
Zohary M 1973 Geobotanical Foundations of the Middle East. Gustav Fischer Verlag, Stuttgart.
תודות
תודה לרינה ולילדי שבילו סופי שבוע רבים בחלקות האלונים. תודה לעוזי פז שבחר עבורי את התמונות מהאוסף הנהדר שלו והתיר את פרסומן.
==================================
כל הזכויות שמורות ל"כלנית" ©
ציטוט: נאמן ג 2016 שונות בפנולוגיה של עלוות אלון התבור: האם אלון התבור נמצא בשלבי מעבר מעץ נשיר חורף לירוק עד? כתב-עת "כלנית" מספר 3.
https://www.kalanit.org.il/quercus-ithaburensis










