ניב דה מלאך, המחלקה לאקולוגיה, אבולוציה והתנהגות, האוניברסיטה העברית בירושלים. nivdemalach@gmail.com
הילה סגרה, הפקולטה לארכיטקטורה ובינוי ערים, הטכניון. hilasegre@gmail.com
מאמר זה מסכם תוצאות מחקרים שבחנו את השפעת רעייה על עושר המינים העשבוני בישראל. המחקרים נערכו בגליל, בגולן, בכרמל, בשפלת יהודה ובנגב וכללו השפעות של רעיית בקר ורעיית צאן. התוצאות מראות שהגורם החשוב ביותר בקביעת התגובה של מין הצמח לרעייה הוא הגובה שלו. רעייה מעלה את השכיחות היחסית של מינים נמוכים כולל גיאופיטים ומינים חד-שנתיים (בעיקר קטניות, מצליבים, ומורכבים), ומורידה את השכיחות היחסית של מינים גבוהים. רעייה מגדילה, במרבית המקרים, את עושר המינים באמצעות מניעת דחיקה תחרותית של מינים נמוכים, ומכאן שרעייה יכולה לשמש ככלי ממשק חשוב לשמירה על מגוון ביולוגי גבוה של צמחים בישראל. למרות זאת, בתנאים מסויימים כמו מפנים דרומיים, הרעייה אינה מעלה את מגוון המינים ובמקומות מסוימים כמו במכלאות ושקתות, הרעייה מביאה להשתלטות של מינים רודראליים ומקטינה בכך את עושר המינים. לכן חשוב להתחשב בתנאי בית הגידול כשקובעים מדיניות ממשק ולהתאים את עצמת ואופי הרעייה ולחציה לתנאים המקומיים.
הקדמה
כמעט כל השטחים בישראל שאינם מעובדים או מיושבים נמצאים תחת רעייה של בקר או צאן כולל שמורות טבע, שטחי אש ויערות נטועים (זליגמן וחובריו, 2016). בעוד שרעייה מאפיינת את אזורנו מזה אלפי שנים, עוצמת הרעייה ואופי הרעייה משתנים לאורך השנים (Seligman and Perevolotsky, 1998). לרעייה שתי השפעות עיקריות: הראשונה והברורה היא אכילה של הצומח והקטנת הביומסה הצמחית. השנייה, המורכבת יותר, היא השפעה על הנוטריינטים בקרקע שעליה נרחיב בהמשך. המאמר מתחיל בהסבר קצר על רעייה בהקשר לתיאוריה האקולוגית אך רובו כולל סקירה של מחקרי הרעייה שהתבצעו בישראל והתמקדו בהשפעות של רעייה על הרכב החברה ועושר מינים של הצמחים העשבוניים. מידע מקיף יותר (השוואות בין ממשקי רעייה שונים, השפעה על צומח מעוצה ועוד הרחבות נוספות) נמצא בספר המרעה החדש הזמין באינטרנט (זליגמן וחובריו, 2016).
רקע
מחקרי המרעה נוהגים לחלק את מיני הצמחים ל'עולים' (increasers), המושפעים לטובה (במונחי שפעה או כיסוי יחסי) מרעייה, ומינים "יורדים" (decreasers) המושפעים לרעה. בעבר היה מקובל שהגורמים העיקריים הקובעים האם מין עולה או יורד כתוצאה מרעייה הם מידת הנעכלות (יחס חלבונים מול סיבים), אחוז המטבולטים המשניים (חומרי הגנה נגד אוכלי עשב, יכולים להיות רעלנים איכותיים או כמותיים) וההגנות המורפולוגיות (למשל קוצים).
מחקר ישראלי, של נוי-מאיר וחובריו, הביא לראשונה ראיות אמפיריות שסתרו את הגישה הזו, והציע השערה אלטרנטיבית שהגורם הראשי הקובע את התגובה לרעייה הוא גובה הצומח (Noy-Meir et al., 1989). לפי השערה זו, בהיעדר רעייה, התחרות על אור מביאה לכך שהמינים הגבוהים מצלים על המינים הנמוכים ומדכאים אותם (DeMalach et al., 2016). לעומת זאת, המינים הגבוהים זמינים יותר לבעלי החיים שלרוב אינם אוכלים בגובה הקרקע, ולכן נפגעים יותר מהרעייה. מאמר סקירה שסיכם תוצאות מחקרים רבים ברחבי העולם, תמך בהשערה של נוי-מאיר וחובריו והראה שגובה הצמח הוא הגורם החשוב ביותר בקביעת התגובה של המין לרעייה (Díaz et al., 2007).
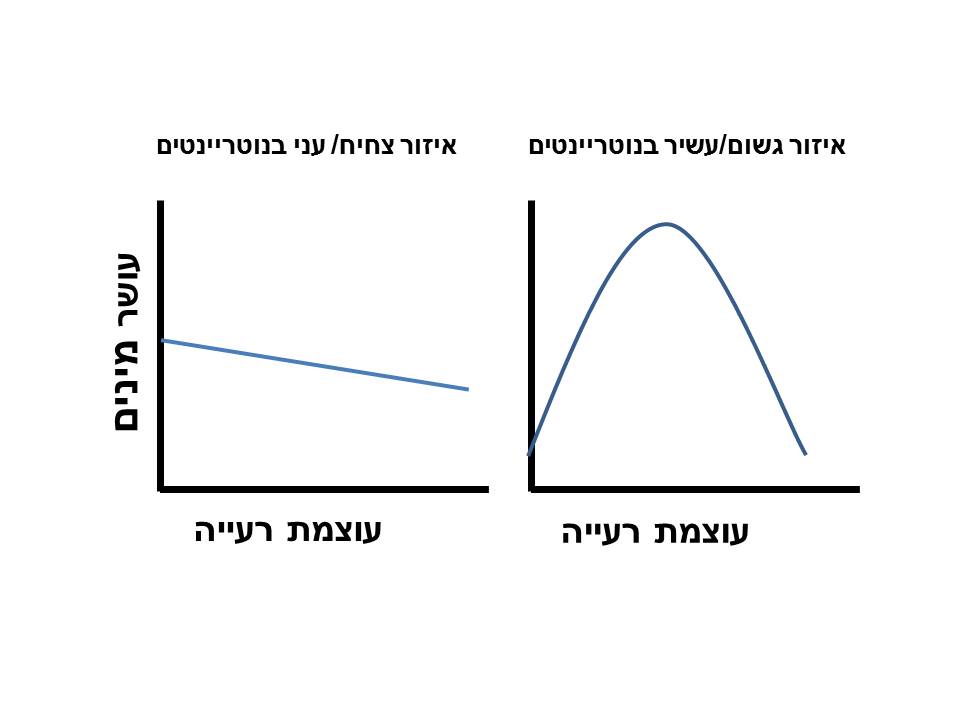
רעייה יכולה להשפיע לא רק על הרכב הצמחייה אלא גם על עושר המינים (מספר המינים ליחידת שטח). לפי המודל הקונספטואלי של מילצ'ונס וחובריו (Milchunas et al., 1988) (איור 1), הנתמך גם על ידי מודלים מתמטיים (May et al., 2009), באזורים מעוטי משאבים (אזורים צחיחים או עניים בנוטריינטים) השפעת רעייה תהיה חלשה אבל בכיוון שלילי. לעומת זאת באזורים בהם זמינות המשאבים גבוהה, השפעת רעייה תהיה אונימודלית (unimodal) – כלומר השפעה חיובית ברמות נמוכות של רעייה, מגוון מקסימלי ברמות בינוניות והשפעה שלילית ברמות גבוהות. המודל מתבסס על ההנחה שגם רעייה וגם מיעוט משאבים יוצרים סלקציה למינים קטנים. לכן, ההשפעה של רעייה על הרכב המינים ועל עושר המינים תחת מיעוט משאבים תהיה קטנה (כי הסלקציה היא באותו כיוון). לעומת זאת, כאשר זמינות המשאבים גבוהה הרעייה מונעת דחיקה תחרותית של מינים נמוכים באמצעות הגדלת זמינות האור (ע"י אכילה) וכך מעלה את עושר המינים. עם זאת, המודל מציע שברמות גבוהות של רעייה עושר המינים יהיה נמוך ללא תלות בתנאי הסביבה כי במקרים כאלו ישרדו רק מינים בעלי עמידות לרעיית יתר.
החיסרון של מודלים תיאורטיים הוא התבססותם על הנחות מופשטות וכלליות ללא התייחסות למורכבות של מערכות טבעיות הכוללת שונות בסוג בע"ח, עיתוי הרעייה, התנאים המקומיים (סוג קרקע, מסלע) הרכב הצומח המקומי והשפעות רעייה על זמינות נוטריינטים (באמצעות הפרשות). בעייה נוספת היא שהמודל מנבא עושר מינים מקסימלי בעצמת רעייה בינונית, אבל קשה להגדיר בדיוק מהי "רעייה בינונית" עבור כל שטח. למרות המגבלות, מחקרים רבים ברחבי העולם תומכים בניבוי של המודל של מילצ'ונס (Milchunas and Lauenroth, 1993; Proulx and Mazumder, 1998; Bakker et al., 2006). לעומת זאת, מאמר סקירה רחב-היקף שסיכם תוצאות ניסויים רבים ברחבי אוסטרליה לא מצא קשר בין תנאי הסביבה לבין השפעת רעייה (Eldridge et al., 2016). בהמשך נציג תוצאות אמפיריות ממחקרים בישראל ונבדוק האם הן מתיישבות עם המודל התיאורטי.
השפעות רעייה על הרכב המינים בישראל
מחקרו החלוצי של נוי מאיר וחובריו (Noy-Meir et al., 1989) שכלל אתרים רבים בצפון הארץ (כרי דשא, הר כנען, שמורת יהודיה) מצא שרעייה גורמת לירידה בשכיחות של מינים גבוהים ועלייה בשכיחות של מינים נמוכים (למעט מין יוצא דופן אחד, עירית גדולה שהיא מין גבוה אבל רעיל ולכן מושפע חיובית מרעייה). כמו כן, נמצאה השפעה למחזור החיים (ירידה בכיסוי הרב-שנתיים), לקבוצה הטקסונומית (עלייה בשכיחות מורכבים) ולמורפולוגיה (עלייה בשכיחות צמחים בעלי שושנת עלים). בניגוד למשוער, המינים הקוצניים (למעט חוח ספרדי), לא היו נפוצים יותר באזורים תחת רעייה.
מחקרים מאוחרים יותר הראו גם הם מגמות דומות. ברמת הנדיב נמצא שרעייה גורמת לעלייה בשכיחות של מינים נמוכים (מתחת ל-15 ס"מ) ומינים המקדימים לפרוח (Hadar et al., 1999). גם בכרי דשא נמצא שרעייה מורידה את הדומינטטיות של הדגנים הגבוהים (שעורת התבור, שיבולת-שועל נפוצה). בנוסף, בניגוד למחקרם של נוי מאיר וחובריו, שטרנברג וחובריו מצאו שהשכיחות של קוצים חד-שנתיים (ממשפחת המורכבים) מצליבים (בעלי הגנות כימיות), עולה בעקבות רעייה (Sternberg et al., 2000; Sternberg et al., 2015). מחקר בלהבים של אוסם וחובריו אשר סיווג את המינים לפי ביומסה במקום גובה, מצא שרעיית צאן פוגעת בעיקר במינים גדולים (שעורת התבור, שיבולת-שועל נפוצה, בן-חיטה ביצני, כליינית מצויה, כרבולת מצויה, אספסת קטועה) ועוזרת למינים קטנים (פילגון מצוי, מיני לחך וניסנית, צללית אשונה, רב-פרי מצוי) (Osem et al., 2004).
קבוצה פונקציונאלית חשובה שעולה בעקבות רעייה היא הגיאופיטים. מחקרים במקומות רבים בגליל ובגולן הראו שששכיחות גיאופיטים גבוהה יותר באזורים תחת רעייה, במיוחד עבור מינים ממשפחות האירוסיים והשושניים[1] (Noy-Meir and Oron, 2001). אותו מחקר מצא, שההשפעה החיובית של רעייה על גיאופיטים עולה ככל שרמת היצרנות עולה. החוקרים שיערו שהסיבה לכך היא שהגיאופיטים נדחקים תחרותית ככל שהביומסה גבוהה יותר והתחרות על אור גדולה יותר. תוצאות מחקר ברמת הנדיב, תמכו גם הם בהשערה זו, נמצא שהפגיעה של רעיית בקר בכלניות היא מינימאלית, בעוד שהורדת הביומסה של המתחרים והגדלת זמינות האור משפיעים לטובה על הכלניות (Perevolotsky et al,. 2011).
דוגמאות להשפעת רעייה על הרכב המינים – ראו בתמונות 2,1.


השפעת רעייה על הרכב מיני צמחים
תמונה 1 (מימין). השוואת הרכב המינים העשבוניים בין חלקות ללא רעייה (לפני הגדר) לחלקות הנתונות לרעייה (מעבר לגדר) בשפלת יהודה (אזור תל גודד). באזור הנתון לרעייה גדלים בעיקר מינים נמוכים (למשל ניסנית דו-קרנית בפריחה צהובה) וגיאופיטים (למשל כלנית מצויה בפריחה אדומה). באזור ללא רעייה גדלים בעיקר מיני דגנים גבוהים (שעורת התבור ושעורת הבולבוסין). צילם: ניב דה מלאך © (פברואר 2012).
תמונה 2 (משמאל). השפעות חיוביות ושליליות של רעייה באזור בית גוברין. האזור ללא רעייה (לפני הגדר הקרובה) מאופיין במגוון נמוך ובשליטה של דגנים, האזור תחת רעייה (מעבר לגדר הקרובה) מאופיין במגוון גבוה ובכיסוי גבוה של רחבי עלים חד שנתיים וגיאופיטים. אזור המכלאה (מעבר לגדר הרחוקה) מאופיין במגוון נמוך ובשליטה של חלמית קטנת פרחים. צילמה: אינגה דירקס © (פברואר 2013).
להגדלה – לחצו על התמונות


השפעות רעייה על עושר המינים בישראל
רוב מחקרי הרעייה בישראל מראים שהשפעת רעייה על עושר המינים של צמחים עשבוניים היא חיובית (Olsvig-Whittaker et al., 1993; Noy-Meir, 1995; Hadar et al., 1999; Sternberg et al., 2000, ;רותם וחובריו, 2015) או נייטרלית (Zaady et al,. 2001; Agra and Ne’eman, 2009) (טבלה 1). ירידה בעושר המינים נמצאה רק בבתי גידול ספציפיים (באף אחד מהמחקרים לא נמצאה ירידה בעושר המינים בכל בתי הגידול). מחקרים שהשוו בתי גידול שונים באותו אתר באזורי המעבר בין האקלים הים התיכוני לאקלים המדברי (להבים ובית גוברין) תמכו בניבוי של המודל של מילצ'ונס שחוזה שהשפעת רעייה תהיה חיובית בעיקר באזורים עשירים במשאבים (Osem et al., 2002; Segre et al., 2016). בבתי גידול פרודוקטיביים (כמו עמקים קטנים שבהם קרקע עמוקה שמעלה את זמינות המים והמינרלים) רעייה העלתה את עושר המינים ולעומת זאת בבתי גידול יובשניים יותר כמו מדרונות המאופיינות בקרקע רדודה, ההשפעה הייתה נייטרלית (בית גוברין) או שלילית (להבים). בניסויי תחרות בבית גוברין נמצא שההבדלים בין המדרון לעמק נובעים משני גורמים שונים הפועלים במקביל: 1. עוצמת הדחיקה התחרותיות בעמק גדולה מאשר במדרון בגלל תחרות א-סימטרית על אור (Segre et al., 2014; דה מלאך, 2017). 2. האפקטיביות של הרעייה בהקטנת הדחיקה התחרותית גדולה יותר בעמק מכיוון שהורדת ביומסה היא אפקטיבית במניעת תחרות על אור אך לא במניעת תחרות על נוטריינטים ומים (Segre et al., 2016).
לעומת זאת כאשר משווים בין מחקרים בדרום למחקרים בצפון, לא נראה שיש הבדל גדול בין השפעת רעייה באזורים גשומים לבין השפעתה באזורים יובשניים (טבלה 1). ממצאים אלו הם בניגוד למודל של מילצ'ונס שניבא קשר בין כמות המשקעים לבין השפעת רעייה. כנראה שגורמים אחרים (מאגר המינים, סוג הקרקע, סוג הרעייה, טמפרטורה) חשובים יותר בקביעת התגובה לרעייה מאשר כמות המשקעים. כמו כן, כמות המשקעים היא לא הגורם היחיד הקובע את זמינות המים, שנקבעת על ידי גורמים נוספים כמו מפנה, נגר, טמפרטורה, וסוג קרקע. בנוסף, חשוב להדגיש שלפי המודל יש חשיבות גדולה לעוצמת הרעייה אך רק שני מחקרים בארץ בדקו את ההשפעה של עוצמות שונות של רעייה. המחקר של נאווה ווויטאקר מצא השפעה אונימודלית, כלומר עושר המינים המקסימלי הופיע בעצמת רעייה בינונית (Naveh and Whittaker, 1980). המחקרים של שטרנברג וחובריו (Sternberg et al., 2000, Sternberg et al., 2015) לא הראו הבדלים בין ההשפעות של רעייה בינונית להשפעות רעייה חזקה.
טבלה 1. סיכום ההשפעה של רעייה על עושר מיני הצמחים העשבוניים, על פי מחקרים שבוצעו בישראל מסודרים מדרום לצפון.
| מקור | אזור בארץ | צפיפות רעייה (בע"ח לקמ"ר) | בית גידול | השפעה |
| Olsvig-Whittaker et al., 1993 | רמת נגב (חולות משאבים) | 150 כבשים (נמוכה), 200 כבשים (בינונית). צפיפות גבוהה לפי הערכה | חיובית | |
| רותם וחובריו 2015 | נגב (ליד ירוחם) | אין נתונים כמותיים | מדרון | חיובית |
| ערוץ נחל | חיובית | |||
| Zaady et al,. 2001 | צפון הנגב (ליד אופקים) | 100 כבשים | מדרון דרומי | נייטרלית |
| מדרון צפוני | נייטרלית | |||
| Osem et al,. 2002 | צפון הנגב (ליד להבים) | 100 כבשים | מדרונות | שלילית |
| כתפי ואדי | חיוביות | |||
| Segre et al., 2016 | שפלת יהודה (בית גוברין) | 30 פרות | מדרון | נייטרלית |
| עמק | חיובית | |||
| Sternberg et al., 2000 | גליל מזרחי (כרי דשא) | 55 פרות (נמוכה),110 פרות (בינונית),220 פרות (גבוהה) | חיובית | |
| Hadar et al., 1999 | רמת הנדיב | 230-260 פרות | חיובית | |
| Naveh and Whittaker, 1980 | גליל עליון ותחתון, כרמל וגלבוע | אין נתונים כמותיים | אונימודלית | |
| Noy-Meir, 1995 | הגליל המזרחי והגולן | 25-80 פרות | חיובית | |
| Agra and Ne’eman, 2009 | הגליל העליון (נחל צבעון) | 30 פרות | מפנה דרומי | נייטרלית |
| מפנה צפוני | נייטרלית |
השפעת רעייה על הקרקע והשפעות עקיפות על הצומח

במערכות מרעה סגורות, ניתן לצפות שרעייה לא תשפיע על זמינות הנוטריינטים בקרקע כי בעלי החיים צורכים את הנוטריינטים מהצמחים אך מחזירים אותם למערכת באמצעות ההפרשות שלהם (תהליך הקרוי מיחזור יסודות). אולם בפועל, מערכות מרעה לעולם אינן סגורות. חלק מהנוטריינטים שנלקחים מהצמחים נטמעים ברקמות בעלי חיים ויוצאים מהשטח, כאשר הפרות, הכבשים או העיזים הופכת לבשר למאכל. בנוסף, בעלי חיים מבויתים מקבלים תוספת הזנה במהלך העונה היבשה, תוספת זו מכילה נוטריינטים רבים (לרוב עשויה גרעינים או זבל עופות) ולכן מכניסה נוטריינטים למערכת. הפרות והכבשים לא תמיד אוכלות ומפרישות באותם האזורים. האכילה מתבצעת בשטחים נרחבים אבל ההפרשות מתרכזות באזורים מועדפים (ליד שקתות, מכלאות, ואזורים מוצלים). במחקר בו בדקנו את הנוטריינטים הזמינים בקרקע באזורי שקתות בשפלת יהודה מצאנו כמויות גבוהות מאוד של חנקן (איור 1) ומינרלים נוספים. לפי התיאוריה האקולוגית, באזורים כמו שקתות בהן קיימת הפרעה חזקה (אכילה ורמיסה) מצד אחד וזמינות משאבים גבוהה מצד שני, צפויים להופיע צמחי מעזבות (רודראלים) המאופיינים בקצב גידול מהיר ויכולת הפצה גבוהה (Grime, 1977) ואכן מצאנו שאזורים אלו נשלטים על ידי חלמית קטנת-פרחים בליווי של סרפד הכדורים וכף-אווז האשפות. מלבד ההשפעה על הרכב המינים, נמצא שהתנאים באזורי השקתות בשפלה, מורידים את עושר המינים (איור 2). מחקרו של אריה וינוגרד שבדק את הדינמיקה של הצומח והקרקע במכלאות צאן לאחר נטישה ביער פלוגות וביער בארי, הראה שריכוזי החנקן יורדים עם השנים (תוך 10-5 שנים) בגלל שטיפה, אולם ריכוזי הזרחן, שהוא פחות מסיס במים, נשארים גבוהים גם לאחר 20-15 שנה (וינוגרד, 2016). לאורך הזמן הרכב הצומח משתנה ממינים שמופצים דרך מערכת העיכול של בעלי החיים (אנדוזאוכוריה) כמו חלמית קטנת-פרחים וכף-אווז האשפות למינים מופצי רוח, כמו גדילן מצוי שהפרי שלו מכיל ציצית שעירה המסייעת להגדלת מרחק ההפצה. לאחר כ-20-15 שנה חברת הצמחים מתאפיינת בדגנים גבוהים שהם מתחרים טובים על אור בתנאים העשירים שנוצרו אך בעלי יכולת התנחלות (קולוניזציה) נמוכה יותר. עם זאת, גם לאחר 20-15 שנה החברה במכלאות הנטושות שונה מחברת הצומח שמחוץ למכלאות המתאפיינת במינים נמוכים כגון מלעניאל מצוי, רחבי עלים חד-שנתיים (מיני לחך, מקור-חסידה מצוי, מיני ניסנית, חלבלוב מצוי, מרגנית השדה) וגיאופיטים (כלנית מצויה, שום משולש וזמזומית מצויה).

דיון ומסקנות
אנשי שדה רבים טוענים שלרעייה יש השפעות שליליות על העושר של מיני הצמחים. לעומת זאת, סיכום מחקרי המרעה מציגים תמונה הפוכה (טבלה 1, ראו גם זליגמן וחובריו, 2016), רעייה מונעת השתלטות של מינים גבוהים, מקטינה את התחרות על האור ומגדילה את עושר המינים. כמו כן, רעייה עוזרת למינים חד-שנתיים שמהווים חלק גדול מהפלורה בישראל ומגדילה את שכיחות הגיאופיטים.
ראוי לשאול מה מקור הפער בין הרושם של אנשי השדה לבין תוצאות המחקרים. הסבר אחד הוא שקיים פער בין "רושם" לבין "רישום". שטח שעבר כיסוח על ידי רעייה, שהורידה את גובה הצמחייה לגובה דשא נראה עצוב ועני בפרחים, אבל אחרי שבועיים יכול להראות יפה מתמיד. לעומת זאת ההשפעה השלילית של דחיקה תחרותית בעקבות מניעת רעייה נראית רק אחרי כמה שנים, ולרוב לא מייחסים אותה להעדר רעייה. יתכן גם שקיימת הטיה בספרות המדעית שכן המחקרים בודקים פחות את הרכב המינים והמגוון שלהם באזורי מכלאות ושקתות, מכיוון שנראה מובן מאליו שבאזור מופר כל כך מגוון המינים נמוך מאוד. כמו כן, יתכן שיש עליה בכמות תוספת ההזנה המוגשת לפרות במרעה ולכן חלקם היחסי של שטחים שהמגוון בהם נפגע עולה בעקבות העליה בכמות הנוטריינטים.
מסקנת המחקר היא שהתיאוריה האקולוגית נותנת קווים כללים לצורך ברעייה (צורך גדול יותר באזורים פרודוקטיביים) אבל ישנם אלמנטים נוספים שהם ספציפיים לבית גידול שונים, ונדרשים כלים שיוכלו לשקלל אלמנטים אלו בחישובי עצמות הרעייה הנדרשות. לשם כך, יש צורך בפיתוח של המודלים התיאורטים לכיוונים ישומיים שיאפשרו להבין מה היחס בין עצמת הרעייה בפועל לבין עצמת הרעייה האופטימלית לשמירת טבע.
מסקנה נוספת היא שרעייה היא כלי ממשקי הכרחי לשמירה על מגוון מינים בישראל, זאת בנוסף לפתיחת החורש וצמצום שרפות. עם זאת, עדיין דרושים מחקרים שייבחנו כיצד מממשקי רעייה שונים (למשל רעיית בקר לעומת צאן או עיזים, השפעת סוג וכמות תוספת ההזנה, עוצמות שונות של רעייה) משפיעים על החלק היחסי של השטח שנפגע , ולשקלל באופן מיטבי את היתרונות והחסרונות של הרעייה במקומות השונים.
תודות
אנו מודים לבר שמש שבזמן דיגום צומח משותף בבית גוברין העלה את הצורך לכתוב מאמר המסכם את השפעות הרעייה בארץ. כמו כן אנו מודים לרשות הטבע והגנים על התמיכה במחקרי הרעייה שלנו בגן לאומי בית גוברין.
ספרות:
דה מלאך נ 2017 היפותזת אסימטריות האור ודחיקה תחרותית של צמחים בעקבות דישון: תוצאות מניסוי בגן לאומי בית גוברין כלנית 4.
וינוגרד א 2016 דינאמיקה של חברת צמחים במכלאות צאן נטושות וסביבתן: תהליכים וממשק (עבודת דוקטורט)
זליגמן נ אונגר י הנקין ז לנדאו י וחובריהם 2016 על צומח, בעלי חיים ואנשים. תורת ניהול המרעה בישראל. הוצאה לאור נקודת ח"ן, ירושלים.
רותם ג אהרון ש רותם נ וקולומבוס א 2015 השפעת רעיית צאן על מבנה חברת צומח באזור מדברי ניתוח תוצאות ראשוניות. תקציר הרצאות כנס האגודה הישראלית למדעי המרעה (תקציר כנס המרעה 2.)
—————————————————————————————————————
Agra H and Ne’eman G 2009 Woody species as landscape modulators: their effect on the herbaceous plants in a Mediterranean maquis. Plant Ecology 205: 165-177.
Bakker ES Ritchie ME Olff H Milchunas DG and Knops JMH 2006 Herbivore impact on grassland plant diversity depends on habitat productivity and herbivore size. Ecology Letters 9: 780-788.
DeMalach NE Zaady J Weiner and Kadmon R 2016 Size asymmetry of resource competition and the structure of plant communities. Journal of Ecology 104: 899-910.
Díaz S Lavorel S McIntyre S Falcuk V et al. 2007 Plant trait responses to grazing – a global synthesis. Global Change Biology 12: 1-29.
Eldridge DJ Poore AGB Ruiz-Colmenero M Letnic M et al. 2016 Ecosystem structure, function, and composition in rangelands are negatively affected by livestock grazing. Ecological Applications 26: 1273-1283.
Grime JP 1977 Evidence for existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. American Naturalist 111: 1169-1194.
Hadar l Noy-Meir I and Perevolotsky A 1999 The effect of shrub clearing and grazing on the composition of a Mediterranean plant community: functional groups versus species. Journal of Vegetation Science 10: 673-682.
May FV Grimm V and Jeltsch F 2009 Reversed effects of grazing on plant diversity: the role of below-ground competition and size symmetry. Oikos 118: 1830-1843.
Milchunas DG and Lauenroth WK 1993 Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecological Monographs 63: 327-366.
Milchunas DG Sala OE and Lauenroth WK 1988 A generalized-model of the effects of grazing by large herbivores on grassland community structure. American Naturalist 132: 87-106.
Naveh Z and Whittaker RH 1980 Structural and floristic diversity of shrublands and woodlands in northern Israel and other Mediterranean areas. Vegetatio 41: 171-190.
Noy-Meir I 1995 Interactive effects of fire and grazing on structure and diversity of Meditterranean grasslands. Journal of Vegetation Science 6: 701-710.
Noy-Meir IM Gutman and Kaplan Y 1989 Responses of Mediterranean Grassland plants to Grazing and Protection. Journal of Ecology 77: 290-310.
Noy-Meir I and Kaplan D 2002 Species richness of annual legumes in relation to grazing in Mediterranean vegetation in northern Israel. Israel Journal of Plant Sciences 50: S95-S109.
Noy-Meir I and Oron T 2001 Effects of grazing on geophytes in Mediterranean vegetation. Journal of Vegetation Science 12: 749-760.
Olsvig-Whittaker LS Hosten PE Marcus I and Shochat E 1993 Influence of grazing on sand field vegetation in the Negev desert. Journal of Arid Environments 24: 81-93.
Osem Y Perevolotsky A and Kigel J 2002 Grazing effect on diversity of annual plant communities in a semi-arid rangeland: interactions with small-scale spatial and temporal variation in primary productivity. Journal of Ecology 90: 936-946.
Osem YA Perevolotsky and Kigel J 2004 Site productivity and plant size explain the response of annual species to grazing exclusion in a Mediterranean semi-arid rangeland. Journal of Ecology 92: 297-309.
Perevolotski AR Schwartz-Tzachor E Yonathan R and Ne’eman G 2011 Geophytes-herbivore interactions: reproduction and population dynamics of Anemone coronaria L. Plant Ecology 212: 563-571.
Proulx M and Mazumder A 1998 Reversal of grazing impact on plant species richness in nutrient-poor vs. nutrient-rich ecosystems. Ecology 79.
Segre H DeMalach N Henkin Z and Kadmon R 2016 Quantifying Competitive Exclusion and Competitive Release in Ecological Communities: A Conceptual Framework and a Case Study. PloS one 11:e0160798.
Segre H Ron R De Malach N Henkin Z et al. 2014 Competitive exclusion, beta diversity, and deterministic vs. stochastic drivers of community assembly. Ecology letters. 17(11): 1400-148.
Seligman NG and Perevolotsky A 1998 Role of Grazing in Mediterranean Rangeland Ecosystems Inversion of a paradigm. BioScience 48.
Sternberg M Golodets C Gutman M Perevolotsky A Ungar E D Kigel J and Henkin Z 2015 Testing the limits of resistance: a 19-year study of Mediterranean grassland response to grazing regimes. Global Change Biology 21: 1939-1950.
Sternberg M Gutman M Perevolotsky A Ungar ED et al. 2000 Vegetation response to grazing management in a Mediterranean herbaceous community: a functional group approach. Journal of Applied Ecology 37: 224-237.
Zaady E Yonatan R Shachak M and Perevolotsky A 2001 The effects of grazing on abiotic and biotic parameters in a semiarid ecosystem: A case study from the Northern Negev Desert, Israel. Arid Land Research and Management 15: 245-261.
הערות
[1] הכוונה לשושניים לפי שיטת המיון הקלאסית. לפי השיטה הטקסונומית המקובלת כיום, משפחה זו פוצלה לכמה משפחות נפרדות.
=======================================
כל הזכויות שמורות ל"כלנית" ©
ציטוט: דה מלאך נ וסגרה ה 2017 השפעת רעייה על עושר והרכב מיני צמחים עשבוניים בישראל – שיפור או דיכוי?, כתב-עת "כלנית" מספר 4.










