מיכל סמוני-בלנק, הפקולטה לביולוגיה בטכניון, מכון טכנולוגי לישראל, קריית הטכניון michal.samuni@gmail.com
עדו יצחקי, החוג לביולוגיה אבולוציונית וסביבתית, אוניברסיטת חיפה izhaki@research.haifa.ac.il
זאב ארד, הפקולטה לביולוגיה בטכניון, מכון טכנולוגי לישראל, קריית הטכניון zarad48@gmail.com
יורם גרשמן, החוג לביולוגיה, מכללת אורנים gerchman@research.haifa.ac.il
תרגם, ערך והביא לדפוס: גידי נאמן, החוג לביולוגיה וסביבה, אוניברסיטת חיפה-אורנים ומערכת כלנית gneeman@gmail.com
מבוסס על המאמרים:
Samuni-Blank M Izhaki Dearing MD Gerchman Y Trabelcy B Lotan A Karasov WH Arad Z 2012 Intraspecific Directed Deterrence by the Mustard Oil Bomb in a Desert Plant. Current Biology 22: 1218- 1220. DOI 10.1016/j.cub.2012.04.051
Samuni-Blank M Arad Z Dearing MD Gerchman Y Karasov WH Izhaki I 2013 Friend or foe? Dispa
plant–animal interactions of two congeneric rodents. Evolutionary Ecology 27:1069-1080. DOI 10.1007/s10682-013-9655-x
Samuni-Blank M Izhaki I Gerchman Y Dearing MD Karasov WH Trabelcy B et al. (2014) Taste and Physiological Responses to Glucosinolates: Seed Predator versus Seed Disperser. PLoS ONE 9(11): https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0112505
תמונת השער: קוצן מצוי, צילמה מיכל סמוני-בלנק ©
תקציר: מים ומזון הם גורמים מגבילים עבור בעלי חיים במדבר. פירות עסיסיים של צמחים הם נדירים במדבר ולכן מושכים מגוון של בעלי חיים הניזונים מהם. הפירות מכילים סוכרים, שומנים וחלבונים כגמול עבור אוכלי הפירות הניזונים מהם ומפיצים את הזרעים. רכפתן מדברי מכיל בציפה של פירותיו המתוקים ובזרעים, מטבוליטים משניים (גלוקוזינולטים). אלה משמשים להגנה על הצמח, ובעיקר על אברי הרבייה שלו, מפני אוכלי צמחים (הרביוורים). במאמר זה אנו מציגים את היחסים ההדדיים בין שני מכרסמים, לבין רכפתן מדברי החולקים אתו בית גידול משותף, וניזונים מפירותיו. קוצן זהוב הוא טורף זרעים, האוכל את הפירות ובמרבית המקרים, גורס את הזרעים בשיניו. לעומתו, קוצן מצוי אוכל את הפירות, יורק את הזרעים הרחק מצמח האם וכך מפיץ אותם. בפירות ובזרעים של הרכפתן מצויה 'פצצת שמן החרדל'. בציפת הפרי נמצא גלוקוזינולט שהוא מר אך אינו רעיל, ובזרעים נמצא האנזים מירוזינאז אשר מפרק את הגלוקוזינטים לחומרים רעילים. קוצן זהוב, אוכל את הפירות ואת הזרעים ולכן חשוף לחומרי הרעל. לעומתו קוצן מצוי אוכל את הפירות ויורק את הזרעים, וכך הוא אינו מושפע מהרעל ומפיץ את זרעי הרכפתן. במאמר אנו מביאים תוצאות של תצפיות בהתנהגות, ושל ניסויים המבהירים את היחסים המורכבים בין שלושת האורגניזמים. לצפייה בסרטון המציג את עיקר התוצאות לחצו על הקישורית.
מבוא
רכפתן מדברי
רכפתן מדברי Ochradenus baccatus הוא שיח שגובהו כמטר, הוא בעל גבעולים רותמיים, ירוקים ומטמיעים. הצמח מלבלב בתחילת החורף ומשיר את עליו בהתאם לכמויות המים בקרקע. בישראל הוא נפוץ, במקומות עם משק מים משופר יחסית, בוואדיות בנגב ובמדבר יהודה. הרכפתן המדברי הוא מין סודני שתפוצתו העיקרית בצפון אפריקה ובכל האזור הסודני בערב, פרס ופקיסטן, ומגיע בישראל לגבול תפוצתו הצפוני. הפרי הוא ענבה בשרנית לבנה בקוטר של 4 -6 מ"מ (איור 1), המכילה מספר זרעים קטנים בקוטר של כמ"מ אחד. הפרי צבעו לבן, הוא מתוק ונאכל על-ידי בעלי חיים רבים, בעיקר צפרים ויונקים (שמידע ואור 1983).
רכפתן מדברי פורח בעיקר באביב, אך הפריחה נמשכת בכמויות משתנות במשך כל השנה. בעבר הוא תואר כצמח דו-ביתי, כלומר שפרטים שונים נושאים פרחים או זכריים, או פרחים נקביים. במחקר שנערך בישראל נמצא שמנגנון הזיווג מורכב יותר. באוכלוסיות הרכפתן המדברי יש פרטים שהם נקבות המייצרות פירות וזרעים, אחרים הם זכרים שמפיצים רק אבקה ואינם מייצרים פירות. מצויים גם פרטים שמרבית פרחיהם זכריים, אך הם נושאים גם פרחים נקביים. 35% מהצמחים הדו-מיניים לא יצרו פירות ו 65% יצרו כמויות משתנות של פירות וזרעים. פרטים גדולים יצרו יותר פירות מאשר פרטים קטנים וצמחים נקביים. צמחים נקביים יצרו פי 20-9 יותר זרעים מאשר צמחים דו-מיניים עם כמויות משתנות של פרחים נקביים, והזרעים שלהם נבטו וצמחו טוב יותר. יצירת פירות על ידי פרטים דו-מיניים ירדה לאורך גרדיינט היובש, ממפנה צפוני לדרומי, כלומר מתנאים טובים לקשים יותר (Wolfe & Shmida 1995, Wolfe & Shmida 1997). במחקר אחר שנערך במצרים, נמצאו שני גלי פריחה, באביב ובסתיו. הפריחה האביבית יצרה יותר פירות וזרעים. ובדומה למצב בישראל, צמחים נקביים יצרו יותר פירות מפרטים דו-מיניים, וצמחים גדולים יצרו יותר פירות מצמחים קטנים (Hegazy et al. 2011). במחקר אחר בישראל, נמצאה השפעה של תנאי בית הגידול על תכונות הפירות והזרעים. רכפתן מדברי שגדל בקרקע עשירה יחסית במים ובחנקן, יצר פירות גדולים יותר שהכילו יותר זרעים, אך פחות שומנים, סוכרים, חנקן ומגנזיום מאשר צמחים שגדלו בקרקע דלה (Lotan and Izhaki 2013).
הפצת זרעים
הזרעים של צמחים מתפתחים מהביציות לאחר הפרייתן. במכוסי הזרע הביציות נמצאות בתוך השחלה ההופכת לפרי. מכיוון שיש יתרונות רבים להפצה של זרעים הרחק מצמח האם שלהם, התפתחו בצמחים התאמות שונות להפצת הזרעים והעיקריים הם על-ידי הרוח (אנמוכוריה) על גבי בעלי-חיים (אפיזואוכוריה) ובתוך בעלי-חיים (אנדוזואוכוריה), אשר אוכלים את הפירות ומפרישים את הזרעים הבלתי פגועים (Pijl 1978). יחידות הפצת הזרעים הן מגוונות בצמחים שונים, אבל חולקות תכונות עיקריות משותפות. על-פי תכונות אלה מקובל לשייך את הצמחים לסינדרומים שונים של ההפצה (Pijl 1978). על מנת שתהיה הפצה יעילה חייבת להיות התאמה בין תכונות הצמח לאלה של בעלי החיים המפיצים את הזרעים. במרבית המקרים, התפתחו התאמות הדדיות אלה בתהליך הנקרא קו-אבולוציה (Herrera 1985). כך למשל, צמחים בעלי הפצה על-ידי אכילה, יש להם פירות צבעוניים שבעלי החיים מגלים בקלות, הם עסיסיים ומכילים חומרים מזינים ובעיקר סוכרים, שומנים ומעט חלבונים.
בישראל (שמידע וארונסון 1983) ובדרום ספרד (Herrera 1995) נמצא כי לשיחים, מטפסים ועצים יש פירות עסיסיים וצבעוניים, המבשילים על הצמחים ומכילים זרעים קטנים, הם נאכלים על פי רוב על-ידי ציפורי שיר הבולעות את הזרעים ומפיצות אותם בלשלשת שלהם. הציפורים נמשכות לפירות בשל ערכם המזוני הכולל בעיקר סוכרים, אך גם שומנים וחלבונים (Dennis 2007).
בצמחיה הים-תיכונית נמצאו התאמות רבות בין ציפורי שיר קטנות הניזונות בעיקר על פירות, לבין תכונות הפירות הקטנים. ההתאמות כוללות את צבע הפירות, שמרביתם אדומים כחולים או שחורים (שמידע וארונסון 1983). התאמה עונתית בין השפע והמגוון של הפירות המבשילים בעיקר בסתיו, לבין לשפע והמגוון של צפורי השיר הנודדות והחורפות, נמצאה במחקרים בישראל ובדרום ספרד (Izhaki et al. 1981, שמידע ואהרונסון 1983Herrera 1995, ). ההתאמות העיקריות שנמצאו הן מורפולוגיות בין מידות המקור לגודל הפירות, אך לא נמצאו הבדלים פסיולוגיים בין צפרים אוכלות פירות לבין אוכלות החרקים, למעט זמן קצר יותר של מעבר המזון דרך המעי (Herera 1995). בעבר סברו חוקרים כי כל המקרים בהן קיימות התאמות הדדיות, מוטואליסתיות, התורמות לכשירות של שני המינים, מקורם בתהליך קואבולוציה. אולם, מסתבר שהצמחים הים תיכוניים בעלי פירות וזרעים מופצי צפורים, נמצאו באגן הים-התיכון כבר בתקופות גאולוגיות קדומות (אאוקן, אוליגוקן מיוקן), הרבה לפני היווצרותו של האקלים הים תיכוני בפליאוקן, ולפני שגובשו מסלולי הנדידה של הצפרים המפיצות את זרעיהם בפלייסטוקן (Herera 1995).
פירות הרכפתן המדברי עשירים בסוכרים (85%, על בסיס משקל יבש) אך דלים בחנקן (2.6%), לעומת זאת הזרעים עשירים בחלבונים (25%) (Samuni-Blank et al. 2012). הפירות הבשלים של הרכפתן המדברי נשארים על הצמח ואינם נושרים, כמו מרבית הפירות מופצי הצפרים. אולם הצבע הלבן של פירות הרכפתן יוצא דופן שכן כמעט כל הפירות האחרים הם צבעוניים (שמידע וארונסון 1983). בישראל נמצא שבולבול ממושקף Pycnonotus xanthopygos, מפיץ זרעים של רכפתן מדברי בעיקר בטווח קצר, בתחום אגן ההיקוות של אותו נחל בו גדלה האם, בעוד שטריסטרמית Onychognathus tristramii, מפיצה את הזרעים למרחקים גדולים ולאגני היקוות של נחלים סמוכים (Spiegel and Nathan 2007).
מכרסמים
בשל ערכם המזוני הגבוה, זרעים מהווים מקור מזון חשוב למינים רבים של בעלי חיים . בניגוד לפירות בשלים הם זמינים, כמעט, במשך כל השנה וניתנים לאגירה (Brown et al. 1979, Vander Wall 1990). מכרסמים ככלל ועכברים בפרט הם גרניוורים, כלומר ניזונים בעיקר על זרעים אותם הם אוספים לפני או לאחר הפצתם מהצמחים (Jansen et al. 2004). הם מצטיינים במציאת זרעים בקרקע (Abramsky 1983). הם מטפסים וחופרים מצטיינים, בעלי שיניים ושרירי לסת המותאמים לאכילת זרעים קשים, הם מכרסמים אותם תוך שהם מחזיקים אותם ברגליהם הקדמיות (Ben-Moshe et al. 2001). לחלקם יש כיסים מיוחדים בלחיים, בהם נאסף המזון (Brown et al. 1979). בשל יעילותם הרבה בטריפת זרעים, למכרסמים יש השפעה ניכרת על הצלחת הרבייה והכשירות של צמחים, על תפוצתם ועל המגוון שלהם (Abramsky 1983, Brown and Heske 1990, Velho et al. 2012). שפע של זרעים יכול להשפיע על הגודל והדינמיקה של אוכלוסיות מכרסמים (Brown et al. 1979). למרות היותם בעיקר טורפי זרעים, למכרסמים יש גם תפקיד בהפצת זרעים, בעיקר על-ידי איסוף ואגירה של זרעים גדולים, לעיתים במקומות המשפרים את הישרדותם ואת סיכויי ההתבססות של הנבטים (Vander Wall 1990). אכילה רשלנית, בה זרעים הולכים לאיבוד תוך כדי אכילה, נפוצה בבתי גידול פוריים, אך נדירה יותר בבתי גידול מוגבלי משאבים כמו מדבריות (Price and Joyner 1997).
רכפתן מדברי, גדל בנחלים במדבר עם שוליים סלעיים, יחד עם כמה מיני מכרסמים וביניהם: קוצן זהוב Acomys russatus שהוא פעיל יום, קוצן מצוי Acomys cahirinus שהוא פעיל לילה, יפה זנב Sekeetamys calurus ונוספים. היחסים ההדדיים בין שני מיני הקוצנים נחקרו רבות בישראל ולא נסקור אותם כאן (Shkolnik 1966, Shkolnik & Borut 1969, Kronfeld-Schor & Dayan 1999, Jones & Dayan 2000, Haim et al. 2005, Gutman et al. 2006 ).
מטבוליטים משניים
מטבוליטים משניים הם חומרים המצויים בצמחים, אבל אינם חלק מתהליכי החיים הבסיסיים ברמת התא. צמחים רבים מכילים מטבוליטים משניים בריכוזים שהם רעילים או דוחים בעלי חוליות ובמיוחד אוכלי זרעים, ולהם השפעה ניכרת על היחסים ההדדיים שבין הצמחים לבעלי החיים (Herrera 1982, Bennett & Wallsgrove 1994, Izhaki 2002, Samuni-Blank et al. 2012, Samuni-Blank et al. 2013, Samuni-Blank et al. 2014).
שמני חרדל הוא השם המקובל לגלוקוזינולטים (Glucosinolates) שהם מטבוליטים משניים הנפוצים בצמחים מסדרת הצלפאים, כולל משפחת המצליבים והרכפתיים. לכשעצמם הם מרים אך בלתי רעילים. לעומת זאת, תוצרי הפירוק שלהם רעילים ונוצרים עקב פעילותו של האנזים מירוזינאז (Myrosinase). הגלוקוזינולטים והמירוזינאז מאוכסנים בצמח בתאים שונים. כאשר רקמת הצמחים נפגעת, התאים משחררים את האנזים המפרק את הגלוקוזינולטים השונים לחומרים (Thiocyanates, isothiocyanates, nitriles) שהם רעילים לבעלי חיים שונים (איור C1) (Halkier & Gershenzon 2006, Winde & Wittstock 2011). מנגנון ביוכימי זה כונה 'פצצת שמן החרדל' (mustard oil bomb) (Matile 1980) והוא נמצא יעיל בהגנה על צמחים בפני הרביוורים ג'נרליסטים (Wittstock & Haliker 2002).
נכוחותם של חומרי הגנה בעלים של צמחים ברורה, אך מדוע הם נמצאים גם בפירות הנאכלים על-ידי בעלי חיים? לפחות שבע תאוריות הוצעו כדי לענות על שאלה זו. יש הטוענים שהחומרים מזרזים את מעבר המזון דרך המעיים, ובכך את הפרשת הזרעים שנבלעו. יש הסוברים שאכילת הפירות משפרת את הבריאות, משך החיים והביצועים של בעלי החיים. מספר תאוריות סוברות שהשפעת החומרים היא שונה בהתאם לבעל החיים האוכל אותם (Cipollini & Levey 1997, Cipollini 2000). כך הראו למשל, שצפרים אוכלות פירות המכילים מטבוליטים משניים כמו פירות הצ'ילי החריף, בעוד שמכרסמים נמנעים מלאכול אותם (Tewksbury & Nabhan 2001, Cipollini & Levey 1997, Levey et al. 2006).
תאוריית "ההרתעה הישירה" גורסת שלמטבוליטים משניים, הנמצאים בפירות בשלים, יש השפעה זניחה על מפיצי הזרעים, אבל הם דוחים טורפי זרעים בשל טעמם הרע או רעילותם. יש רק מעט נתונים התומכים בתאוריה זו, ובמיוחד לגבי בעלי חיים קרובים זה לזה, שאינם משוייכים למחלקות שונות כמו ציפורים ומכרסמים. במאמר זה, אנו בודקים באופן ישיר את התאוריה בעיקר לגבי שני מיני מכרסמים קוצן מצוי וקוצן זהוב, החיים יחדיו בבית הגידול של רכפתן המדברי וניזונים מפירותיו. באופן מיוחד, נתייחס להבדלים בהשפעות הפיסיולוגיות וההתנהגותיות של פצצת שמן החרדל עליהם, ונבחן באיזו מידה הם טורפים או מפיצים של זרעי הרכפתן.
תוצאות ודיון
רכפתן מדברי
נמצאו ריכוזים גבוהים של גלוקוזינולטים בשורשים, בגבעולים, בעלים ובציפת הפרי של רכפתן מדברי. נמצאה גם הפרדה ברורה של מרכיבי פצצת שמני החרדל כאשר הגלוקזינולטים נמצאים בציפת הפרי והאנזים המשפעל מירוזינאז נמצא בזרעים (איור 1A,B). נזכיר שהגלוקוזינולטים אינם רעילים, ורק פירוקם על ידי האנזים הופכת אותם לרעילים. לכן, כל האוכל את הפירות המתוקים ובולע את הזרעים הקטנים בשלמותם מבלי לפגוע בקליפתם, אינו מושפע מהרעילות. כך גם הדבר בבני האדם הנוהגים לאכול את פירות הרכפתן המתוקים.
 איור 1. הפרדה בין הגלוקוזינולטים הנמצאים בציפת הפרי (A), לבין האנזים מירוזינאז, המפרק את הגלוקוזינולטים
איור 1. הפרדה בין הגלוקוזינולטים הנמצאים בציפת הפרי (A), לבין האנזים מירוזינאז, המפרק את הגלוקוזינולטים
לחומרם רעילים (B) והמבנה הכימי שלהם (C) (Samuni-Blank et al. 2012).

קוצן מצוי
בצילומים נחשפו מספר פרטים של קוצן מצוי הנחשב לטורף זרעים, כשהם מטפסים בלילה על שיחי רכפתן מדברי, קוטפים צברי פירות, מרחיקים אותם מצמחי האם, אוכלים את הפירות במסתור בין הסלעים ויורקים את הזרעים (סרטון 1S). צילום מקרוב בשביה, הראה דגם התנהגות דומה (סרטון 2S).
גם פרטים של קוצן זהוב (איור B2) ( סרטון 3S), ופרטים של יפה זנב (איור 2C) צולמו כשהם יורקים זרעים לאחר שאכלו פירות של רכפתן מדברי, כלומר התנהגות זו אינה בלעדית לקוצן מצוי (איור A2) (Samuni-Blank et al. 2012).

איור 2. שלושה מיני מכרסמים: קוצן מצוי, קוצן זהוב ויפה זנב יורקים זרעים של רכפתן מדברי לאחר שאכלו את פירותיו
(Samuni-Blank et al. 2012).
פירות שהונחו מתחת לשיחי רכפתן מדברי נלקחו, נאכלו במסתור בין אבנים ושם נמצאו הזרעים. מסתורים מוצלים כאלה, הם אתרים המגדילים את ההסתברות לנביטה והתבססות של רכפתן מדברי. כל הזרעים שנאספו בשדה לאחר שקוצן מצוי ירק אותם הונבטו במעבדה. 74% מפירות רכפתן שהוצעו לקוצנים מצויים נאכלו על-ידם. 83% מהזרעים שהם ירקו נבטו, בהשואה ל 90% נביטה של זרעים שהופרדו ידנית מהפירות, וההבדל היה מובהק (Z = 21.32, p = 0.18) (Samuni-Blank et al. 2012). אחוז נביטה זה, גבוה פי שניים ממה שנמצא במחקר קודם עבור זרעים שהוצאו מפירות שלא נאכלו (Spiegel & Nathan 2007, Spiegel & Nathan 2011). כאשר קוצנים מצויים הואכלו במעבדה בפירות שלמים, שעברו טיפול לנטרול פעולת האנזים מירוזינאז, הם ירקו רק 20% מהזרעים, יחסית ל 73% מהזרעים בפירות שלא טופלו, והבדל זה היה מובהק (Mann-Whitney U = 46.5, p < 0.001) (Samuni-Blank et al. 2012).
בניסוי נוסף נבדק השינוי במסת הגוף של שמונה קוצנים מצויים לאחר שהואכלו במשך ארבעה ימים בדיאטות שונות: 1. תערובת מעוכה של פירות וזרעים. 2. פירות עם זרעים בלתי מעוכים. 3. תערובת מעוכה שבה נוטרלה פעילות האנזים מירוזינאז. 4. רק זרעים, 5. פירות ללא זרעים (איור 3). רק בטיפול אחד ושניים היו כל המרכיבים הדרושים להפעלת פצצת שמני החרדל, ובהתאם הם גרמו לירידה במשקל הקוצנים, אם כי הירידה בטיפול השני לא הייתה מובהקת. בכל שאר הטיפולים בהם היה חסר מרכיב אחד משני המרכיבים הדרושים להפעלתה, שמרו הקוצנים על משקל גופם במשך ארבעת ימי הניסוי (איור 3) (Samuni-Blank et al. 2012).
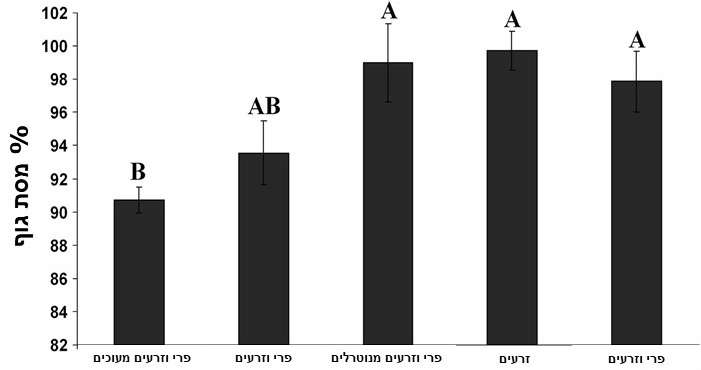
איור 3. השנוי היחסי במסת הגוף של קוצנים מצויים לאחר ארבעה ימים בהם הואכלו בדיאטות שונות:
1. תערובת מעוכה של פירות וזרעים (Activated Mash). 2. פירות עם זרעים בלתי מעוכים (GLSs & Seeds).
3. תערובת מעוכה שבה נוטרלה פעילות האנזים מירוזינאז (Deactivated Mash).
4. רק זרעים (Seeds) 5. פירות ללא זרעים (GLSs).
עמודות המסומנות באותיות זהות אינן נבדלות באופן מובהק. (Samuni-Blank et al. 2012).
כאמור, פצצת שמן חרדל היא מנגנון הגנה מוכר היטב של צמחים בפני בעלי חיים הרביוורים. התוצאות כאן מראות לראשונה תפקיד חדש של המנגנון הגורם למכרסם שהוא בעיקר טורף זרעים להפוך למפיץ שלהם. נמצא שקוצן מצוי נמנע מהשפעה של פצצת שמן חרדל בכך שהוא אוכל את הפירות, אך יורק את הזרעים בהם מצוי האנזים מירוזינאז המפרק את הגלוקוזינולטים לחומרים רעלים. ברגע שבאופן ניסויי עוכבה פעולת האנזים, חזר הקוצן המצוי להתנהגותו הרגילה כטורף זרעים. הזנה בשבי של קוצנים במזון שהכיל את שני מרכיבי פצצת שמן החרדל, גרמה לירידה במשקלם כבר אחרי ארבעה ימים.
קוצן מצוי וקוצן זהוב
בהשוואה שנערכה בין קוצן מצוי וקוצן זהוב, במשך שמונה ימים בשתי אוכלוסיות בשדה, נמצא שבממוצע (±שגיאת תקן) 91.4±8.6% מהפרטים של קוצן זהוב אכלו פירות של רכפתן יחד עם הזרעים, ורק 11.5±5.7% מהפרטים של קוצן מצוי עשו כך, הנותרים ירקו את הזרעים וההבדל היה מובהק (Mann–Whitney U=28, P<0.001) (Samuni-Blank et al. 2013) (איור 4). בניסוי מקביל במעבדה התקבלו תוצאות דומות. תצפיות מקרוב הראו שקוצן מצוי אכל את הפרי וירק את הזרעים, בעוד שקוצן זהוב לעס היטב את הפרי וגרס בשיניו את הזרעים (Samuni-Blank et al. 2013).
קוצן מצוי אכל בממוצע (±שגיאת תקן)0.28±0.01 גר' פירות של רכפתן מדברי בכל יום. קוצן זהוב אכל0.15±0.02 גר' בכל יום, וההבדל היה מובהק (ANCOVA: F1,13=6.7, P<0.05). המספר הממוצע (±שגיאת תקן) של הזרעים השלמים שהושארו על-ידי קוצן מצוי היה 27.5±6.1. קוצן זהוב השאיר רק 5.8±1.1 זרעים וההבדל היה מובהק (t14=2.47, P<0.05) (Samuni-Blank et al. 2013).

איור 4. אחוז ממוצע (±שגיאת תקן) של הקוצנים שאכלו פירות שלמים כולל הזרעים, או רק את הציפה וירקו את הזרעים,
ואחוז ממוצע (±שגיאת תקן) של זרעים שלמים ושל זרעים חיים שהושארו על ידי קוצן מצוי (עמודות אפורות)
וקוצן זהוב (עמודות לבנות). *** מציין הבדל מובהק בין המינים (P<0.05) (Samuni-Blank et al. 2013).
אחוז הנביטה הממוצע (±שגיאת תקן) של הזרעים מתוך כל הפירות שנאכלו היה 74.4±9.0% בקוצן מצוי, רק 4.0±3.1%; בקוצן זהוב, וההבדל היה מובהק (t19=7.6, P<0.001). בבדיקה כמעט ולא נמצאו זרעים שלמים בגללים של שני המינים, אבל 65% מבין הזרעים שכן נמצאו בשני המינים היו ברי נביטה (Samuni-Blank et al. 2013).
בניסוי מעבדה הואכלו שני מיני הקוצנים בשתי דיאטות: 1. מזון עכברים בתוספת פירות מעוכים של רכפתן מדברי, בהם עורבב תוכן הציפה עם תוכן הזרעים והופעלה פצצת שמן החרדל. 2.מזון עכברים בתוספת פירות מעוכים עם זרעים שבהם עוכבה פעולת האנזים מירוזינאז המפעיל את פצצת שמן החרדל. בכל אחת מהדיאטות, קוצן מצוי איבד 10% ממסת גופו יותר מאשר קוצן זהוב. שני המינים איבדו יותר משקל בדיאטה של הפירות הטבעיים שנמעכו יחד עם הזרעים, ללא הבדל בין המינים (two-way ANOVA: Species: F1,28=22.8, P<0.0001, Diet: F1,28=16.9, P<0.0001 ). המסקנה העולה מהתוצאות היא שפצצת שמן החרדל גורמת לירידה במשקל המכרסמים, באופן ישיר, או בשל הירידה בכמות המזון שהם אכלו, יחסית לכמות מזונם לפני תחילת הניסוי (Samuni-Blank et al. 2013).
בניסוי מעבדה נוסף, נבדקו התגובות של קוצן מצוי וקוצן זהוב, החולקים אותו בית הגידול עם רכפתן מדברי, בהשואה להתגובות של עכבר בית (Mus musculus). עכבר הבית אינו מותאם לחיי מדבר ואינו חשוף לפירות רכפתן מדברי בטבע. לכן, הוא לא אמור להיות מותאם לאתגרים הכרוכים באכילת פירות הצמח. לושת המינים הואכלו בשלוש דיאטות: 1. ביקורת: מזון עכברים. 2. ציפת פירות: מזון עכברים שכלל 80% ציפת פירות של רכפתן מדברי ללא זרעים. כלומר, כוללים גלוקוזינולטים אך לא את תוצרי פצצת שמן החרדל . 3. פירות וזרעים מעוכים: מזון עכברים שכלל 80% פירות וזרעים מעוכים. כלומר כולל את התוצרים הרעילים של פצצת החרדל (איור 5).
משקל גוף: נמצאו השפעות מובהקות של הדיאטות (F2,54 = 33.2, P<0.0001) ושל המינים (F2,54 = 36.9, P<0.0001) על משקל הגוף, אולם כל המינים חזרו למשקל גופם התקין. רק עכבר הבית איבד ממשקל גופו בשתי הדיאטות כבר לאחר 24 שעות. שני מיני הקוצנים שמרו על משקל גופם בדיאטות הביקורת והציפה, קוצן מצוי איבד ממשקל גופו בדיאטת הפירות והזרעים המעוכים (איור A5) (Samuni-Blank et al. 2013).
אכילה: נמצאו השפעות מובהקות של הדיאטות השונות על כמות המזון שנאכלה יחסית למשקל הגוף (F2,54 = 35.5, P<0.0001). לא היה הבדל בין מיני המכרסמים (F2,54 = 2.7, P=0.08), אבל הייתה אינטראקציה מובהקת כלומר, הדיאטות השפיעו בצורה שונה על המינים השונים (F4,54 = 9.2, P<0.0001). קוצן זהוב וקוצן מצוי המשיכו לאכול כרגיל, גם בדיאטה המבוססת על ציפת פרי אשר דיכאה אכילה בעכבר בית. אכילת פירות וזרעים מעוכים דיכאה את האכילה של כל המינים (איור B5) (Samuni-Blank et al. 2013).
עיכול: נמצאה השפעה מובהקת של המינים על העיכול (F2,49 =6.4, P<0.005) אך לא נמצאה השפעה של הדיאטות (F2,49 = 1.6, NS), אבל הייתה אינטראקציה מובהקת כלומר, הדיאטות השפיעו בצורה שונה על המינים השונים (F3,49 = 3.0, P<0.05). לא נמצאה ירידה ברמת העיכול אצל קוצן מצוי בדיאטה של פירות ושל פירות וזרעים מעוכים, קוצן זהוב עיכל פחות ועכבר בית לא עיכל כלל את המזון עם הפירות והזרעים המעוכים (איור C5) (Samuni-Blank et al. 2013). מתוצאות אלה ניתן להסיק שקוצן זהוב טורף הזרעים, פחות רגיש לטעם המר של הגלוקוזינולטים המצויים בציפת פרי הרכפתן, וגם עמיד לחומרי הרעל של פצצת שמן החרדל. הוא היה המין היחיד שלמרות שאכל ועיכל פחות מאשר בביקורת (הבדל לא מובהק), שמר על מסת גופו בשתי הדיאטות שהכילו ציפת פרי וציפת פרי וזרעים מעוכים. קוצן מצוי, מפיץ הזרעים, אכל פחות פירות עם זרעים מעוכים, לא הייתה השפעה על מידת עיכול המזון אך ירד במסת גופו. עכבר הבית, אכל מעט מאוד פירות עם זרעים מעוכים, לא עיכל את המזון וירד במשקלו. התוצאות לגבי שני מיני הקוצנים, תואמות את אלה בניסוי קודם (איור 3). בנוסף לכך, משך שהיית המזון במעי של קוצן זהוב היה ארוך יותר מאשר אצל קוצן מצוי בדיאטה של פירות וזרעים מעוכים, ולכן גם זמן פעילות ארוך יותר לאנזימים מפרקי רעלים (Samuni-Blank et al. 2013).
 איור 5. השפעת האכלה של קוצן מצוי (מפיץ זרעי רכפתן, אחרי יומיים), קוצן זהוב (טורף זרעי רכפתן מדברי, אחרי יומיים)
איור 5. השפעת האכלה של קוצן מצוי (מפיץ זרעי רכפתן, אחרי יומיים), קוצן זהוב (טורף זרעי רכפתן מדברי, אחרי יומיים)
ועכבר בית (לא חשוף לצמחי רכפתן מדברי, אחרי יום) לשלוש דיאטות: 1. ביקורת (עמודות לבנות), 2. ציפת פירות ללא זרעים
(עמודת אפורות), 3. ציפת פירות וזרעים על: מסת הגוף כאחוז ממסת הגוף לפני הניסוי (A), אכילת מזון כאחוז ממסת הגוף
לפני הניסוי (B) ורמת עיכול המזון כאחוז ממסת הגוף לפני הניסוי (C). העמודות מציינות ממוצעים ± שגיאת תקן. עמודות
המסומנות באותיות זהות, בכל מין, אינן נבדלות באופן מובהק (Bonferroni Multiple Comparison, P<0.05)
(Samuni-Blank et al. 2013).

בשל דגם הפעילות היומית שלו, קוצן זהוב חשוף ליותר לקרינת שמש, טמפרטורות גבוהות ופחות אפשרויות תזונה מאשר בן סוגו קוצן מצוי. קוצן זהוב מצויד בהתאמות שונות לתנאי המדבר הקיצוני (Haim et al. 1994), בעוד שחלק מהם אינן נמצאות בקוצן מצוי (Shkolnik and Borut 1969, Kronfeld et al. 1994). קוצן זהוב אוסף בפיו הרבה פירות של רכפתן מדברי לפני שהוא גורס אותם, יתכן שהגורם לכך היא פעילותו היומית והתחרות הקשה על מזון. יתכן גם, שהוא גם עמיד יותר להשפעת פצצת שמן החרדל מאשר בן סוגו הקוצן המצוי. הדבר משתקף בעובדה שהוא שמר על משקל גופו בזמן דיאטה שהכילה את תוצרי פצצת שמן החרדל טוב יותר מאשר קוצן מצוי. יתכן שזה הבסיס העיקר להבדל בהתנהגות שיחור המזון בין שני המינים (Samuni-Blank et al. 2013).
סיכום
'פצצת שמן החרדל' היא מנגנון הגנה מוכר מאוד למניעת נזקים מהרביוורים. היא מורכבת מגלוקוזינולטים שהם מרים אך לא רעילים, והאנזים מירוזינאז המפרק והופך אותם לחומרים רעילים. במרבית הצמחים יש הפרדה בין תאים המכילים גלוקוזינולטים לבין תאים המכילים את והאנזים מירוזינאז, לכן פרוק הגלוקוזינולטים לחומרי רעל, מתרחש רק לאחר פגיעה ברקמות הצמחיות, כששני המרכיבים מתערבבים (Kissen et al. 2009). ברכפתן מדברי מצויה הפרדה מיוחדת בה גלוקוזינולטים מצויים בציפת הפרי ואילו האנזים מירוזינאז נמצא בזרעים. הפרדה זו גורמת לקוצן מצוי, אוכל זרעים בעיקרו, לעקוף את פצצת שמן החרדל על-ידי יריקת הזרעים, ובכך להפוך למפיץ זרעים שאינו נפגע מפצצת שמן החרדל. כאשר ,באופן ניסויי, נוטרלה פצצת שמן החרדל מהזרעים, הוא לא ירק אותם וחזר להתנהג כטורף זרעים. כש קוצן מצוי הואכל במזון המכיל תערובת של שני מרכיבי הפצצה, הוא הושפע באופן שלילי, אבל לא כאשר הוגש לו מזון שהכיל רק מרכיב אחד, או רק השני. השינוי הזה בהתנהגות שיחור המזון הוא מידי, ומדגים את הגמישות של קוצן מצוי להפוך מטורף זרעים למפיץ שלהם בסקלת זמן אקולוגית. לשינוי כזה יש השפעה ישירה על כשירות רכפתן מדברי. בשל היותו מין מפתח בבית הגידול המדברי יש לכך השפעה גם על כלל המערכת האקולוגית. תוצאות המחקר מאשרות את בחינתה של תאוריית 'ההרתעה הישירה' הטוענת שמטבוליטים משניים פועלים על טורפי זרעים, אך יש להם השפעה זניחה על מפיצי זרעים (Cipollini and Levey 1997).התוצאות תומכות בתאוריה בכך שלאכילת פירות עם זרעים, המפעילה את פצצת שמן החרדל ומשחררת חומרים רעילים, היו השפעות שליליות בעיקר על עכבר הבית אך גם על קוצן מצוי, שהם בעיקרם טורפי זרעים. אבל הן אינן תומכות בתיאוריה בכך שלמרות שהקוצן הזהוב הוא טורף זרעים, הוא לא נרתע מלאכול את הפירות עם הזרעים, ובעשותו כך לא ירד במשקלו. לעומת זאת קוצן מצוי, מפיץ הזרעים ירד במשקלו כאשר אולץ לאכול את תוכן הזרעים.
המציאות בה שני מינים בני אותו סוג החיים בבית גידול משותף עם צמח שונים זה מזה בהתנהגות שיחור המזון שלהם היא יוצאת דופן. היחסים בין רכפתן מדברי לשני מיני הקוצנים מורכבת. כאמור הקוצן המצוי, הפעיל בלילה, מהווה מפיץ זרעים היורק אותם הרחק מצמח האם, בעוד בן סוגו הפעיל ביום, הקוצן הזהוב ,הוא טורף זרעים. התוצאות מראות שהקוצן הזהוב, האוכל את הפירות וגורס את הזרעים בשיניו, הוא בעל עמידות טובה יותר להשפעה של פצצת שמן החרדל יחסית לקוצן המצוי, ויחסית לעכבר הבית שאינו פוגש פירות של רכפתן בטבע, ובתנאי הניסוי אכל פחות וירד במשקל גופו יותר מאשר שני מיני הקוצנים. העמידות היחסית של הקוצן הזהוב לפצצת שמן החרדל, נובעת כנראה מלחץ התחרות עם הקוצן המצוי בסביבה המדברית דלת המשאבים בה הם חיים.
ספרות
שמידע א ואור י 1983. הצמחיה הסודנית בישראל. רתם 8.
שמידע א וארונסון א 1983 פירות בר עסיסיים בצמחיית ארץ-ישראל והתאמתם להפצה על ידי בעלי-חיים. רתם 10: 44-5.
Abramsky Z 1983 Experiments on seed predation by rodents and ants in the Israeli desert. Oecologia 7:328–332
Ben-Moshe A Dayan & T Simberloff D (2001) Convergence in morphological patterns and commun organization between Old and New World rodent guilds. American Naturalist Nat 158:484–495.
Bennett RN & Wallsgrove RM 1994 Secondary metabolites in plant defense mechanisms. New phytologist 127: 617-633.
Dennis AJ (Ed) 2007 Seed dispersal: theory and its application in a changing world. CABI.
Brown JH & Heske EJ 1990 Control of a desert-grassland transition by a keystone rodent guild. Science 250:1705-1707.
Brown JH Reichman OJ & Davidson DW 1979 Granivory in desert ecosystems. Annual Review of Ecology and Systematics 10:201-227.
Cipollini M 2000 Secondary metabolites of vertebrate-dispersed fruits: evidence for adaptive functions. Review Chiliensis Historia Naturalis 73: 421-440.
Cipollini ML & Levey DJ 1997 Secondary metabolites of fleshy vertebrate-dispersed fruits: adaptive hypotheses and implications for seed dispersal. American Naturalist 150:346-372.
Gutman R Choshniak I & Kronfeld-Schor N 2006 Defending body mass during food restriction in Acomys russatus: a desert rodent that does not store food. American Journal of Physiology 290:881-891.
Haim A Alma A & Neuman A 2005 Body mass is a thermoregulatory adaptation of diurnal rodents to the desert environment. Journal of Thermal Biology 31:168-171.
Haim A Yedidia I Haim & D Zisapel N 1994 Photoperiodicity in daily rhythms of body temperature, food and energy intake of the golden spiny mouse (Acomys russatus). Israel Journal of Zoology 40:145-150.
Halkier BA &Gershenzon J 2006 Biology and biochemistry of glucosinolates. Annual Review of. Plant Biology. 57: 303-333.
Hegazy AK Lovett-Doust J & El Adawy HA 2011 Size-and season-related sex expression and reproductive performance in gynodioecious Ochradenus baccatus Delile (Resedaceae), at Wadi Degla, Egypt. Flora-Morphology, Distribution, Functional Ecology of Plants 206: 1002-1011.
Herrera CM 1982 Defense of ripe fruit from pests: its significance in relation to plant-disperser interactions. American Naturalist 120:218-241.
Herrera CM 1985 Determinants of plant-animal coevolution: the case of mutualistic dispersal of seeds by vertebrates. Oikos 44:132-141.
Herrera CM 1995 Plant-vertebrate seed dispersal systems in the Mediterranean: ecological, evolutionary, and historical determinants. Annual Review of Ecology and Systematics 26:705-727.
Izhaki I Walton PB & Safriel UN 1991 Seed shadows generated by frugivorous birds in an eastern Mediterranean scrub. The Journal of Ecology 79:575-590.
Izhaki I 2002 Emodin—a secondary metabolite with multiple ecological functions in higher plants (a
review). New Phytologist 155:205-217.
Jansen PA Bongers F & Hemerik L 2004 Seed mass and mast seeding enhance dispersal by a neotropical scatter-hoarding rodent. Ecological Monographs 74:569-589.
Kissen R Rossiter JT & Bones AM 2009 The ‘mustard oil bomb’: not so easy to assemble?! Localization, expression and distribution of the components of the myrosinase enzyme system. Phytochemical Review 8:69-86.
Kronfeld N Dayan T Zisapel N & Haim A 1994 Coexisting population of Acomys cahirinus and A. russatus: a preliminary report. Israel Journal of Zoology 40:177-183.
Kronfeld-Schor N Dayan T 1999 The dietary basis for temporal partitioning: food habits of coexisting Acomys species. Oecologia 121:123-128.
Levey DJ Tewksbury JJ Cipollini ML & Carlo TA 2006 A field test of the directed deterrence hypothesis in two species of wild chili. Oecologia. 150 61-68.
Lotan A & Izhaki I 2013 Could abiotic environment shape fleshy fruit traits? A field study of the desert shrub Ochradenus baccatus. Journal of Arid Environments. 92: 34-41.
Matile PH 1980 The mustard oil bomb. Compartmentation of the myrosinase system. Biochemie und Physiologie der Pflanzen 175:722-731.
Price MV & Joyner JW 1997 What resources are available to desert granivores: seed rain or soil seed bank? Ecology 78:764-773.
Samuni-Blank M Izhaki I Dearing MD Gerchman Y et al. 2012 Intraspecific directed deterrence by the mustard oil bomb in a desert plant. Current Biology 22:1218-1220.
Samuni-Blank M Arad Z Dearing MD Gerchman et al. 2013. Friend or foe? Disparate plant–animal interactions of two congeneric rodents. Evolutionary Ecology 27:1069-1080.
Samuni-Blank M Izhaki I Gerchman Y Dearing MD et al. 2014 Taste and Physiological Responses to Glucosinolates: Seed Predator versus Seed Disperser. PLoS ONE 9(11).
Shkolnik A 1966 Studies in the comparative biology of Israel’s two species of spiny mice (genus Acomys). Department of Zoology, Hebrew University, Jerusalem, Israel, PhD dissertation/
Shkolnik A & Borut A 1969 Temperature and water relations in two species of spiny mice (Acomys). American Society of Mammalogy 50:245-255.
Spiegel O & Nathan R 2007 Incorporating dispersal distance into the disperser effectiveness framework: frugivorous birds provide complementary dispersal to plants in a patchy environment. Ecological Letters 10:718-728.
Spiegel O & Nathan R 2011 Empirical evaluation of directed dispersal and density-dependent effects across successive recruitment phases. Journal of Ecology 100: 392-404.
Tewksbury JJ & Nabhan GP 2001 Seed dispersal. Directed deterrence by capsaicin in chilies. Nature 412:403–404.
Vander Wall SB 1990 Food hoarding in animals. University of Chicago Press, Chicago.
Velho N Isvaran K & Datta A 2012 Rodent seed predation: effects on seed survival, recruitment, abundance, and dispersion of bird-dispersed tropical trees. Oecologia 169:995-1004.
Winde I & Wittstock U 2011 Insect herbivore counteradaptations to the plant glucosinolate-myrosinase system. Phytochemistry 72: 1566-1575.
Wittstock U & Haliker BA 2002 Glucosinolate research in the Arabidopsis era. Trends in Plant Science 7:263-270.
Wolfe LM & Shmida A 1995 Regulation of gender and flowering behavior in a sexually dimorphic desert shrub (Ochradenus baccatus Delile [Resedaceae]). Israel Journal of Plant Sciences, 43:325-337.
Wolfe LM & Shmida A 1997 The ecology of sex expression in a gynodioecious Israeli desert shrub (Ochradenus baccatus). Ecology 78:101-110.
כל הזכויות שמורות ל"כלנית" ©
לציטוט: סמוני-בלנק מ. 2021 "פצצת שמן החרדל" בפירות וזרעים של רכפתן מדברי, והשפעתה על שני מיני קוצנים: קוצן זהוב טורף זרעים, לעומת קוצן מצוי המפיץ אותם. כתב-עת "כלנית", מספר 7.











לפוסט הזה יש תגובה אחת
מעניין ממש. תודה